|
Robert A. Freitas Jr., Xenology:
An Introduction to the Scientific Study of Extraterrestrial Life,
Intelligence, and Civilization, First Edition, Xenology Research
Institute, Sacramento, CA, 1979;
http://www.xenology.info/Xeno.htm
(c) 1979 Robert A. Freitas Jr.
All Rights Reserved. |
Chapter 11.
Extraterrestrial Biomechanics
"...but why must they be of the same nature as ours?
Nature seems to court variety in her Works, and may have made them widely different
from ours either in their matter or manner of Growth, in their outward Shape,
or in their inward Contexture; she may have made them such as neither our Understanding
nor imagination can conceive."
-- Christian Huygens, in The
Celestial Worlds Discover’d; Or, Conjectures Concerning the Inhabitants,
Plants and Productions of the Worlds in the Planets (1698)602
"I would even go so far as to say that, from the cosmic
viewpoint, all terrestrial mammals are ‘humanoid.’ They all have
four limbs, two eyes, two ears, one mouth, arranged symmetrically about a single
axis. Could a visitor from Sirius really tell the difference between a man and,
for example, a bear? (‘I’m terribly sorry, Mr. Prime Minister, but
all humanoids look the same to me....’)"
-- Arthur C. Clarke (1968)373
"Life elsewhere is likely to consist of odd combinations
of familiar bits."
-- Allen Broms, in
Our
Emerging Universe (1961)1191
"Two large dark-coloured eyes were regarding me steadfastly.
The mass that framed them, the head of the thing, it was rounded, and had, one
might say, a face. There was a mouth under the eyes, the lipless brim of which
quivered and panted, and dropped saliva. The whole creature heaved and pulsated
convulsively. A lank tentacular appendage gripped the edge of the cylinder,
another swayed in the air. Those who have never seen a living Martian can scarcely
imagine the strange horror of its appearance...
-- H. G. Wells, in The
War of the Worlds (1898)
The rich diversity of multicellular life on our own planet
should tip us off that nature will have produced similarly intricate designs
elsewhere in our Galaxy. The flora and fauna of extrasolar worlds doubtless
will differ markedly from terrestrial species. One has only to consider the
tremendous variety of terran life to realize at once that we live in a grand
planetary zoo without equal -- save that of another world. Nevertheless, there
must exist some limits to strangeness, some basic, universal rules regarding
the construction of physical beings.
Our imagination sorely needs stimulation rather than restraint.
So perhaps it is appropriate to begin by attacking one of our most basic assumptions
about alien life: That all organisms must be built of cells. Since phase separation
is probably a fundamental prerequisite to life, and since small cells are virtually
certain to arise prebiotically, it was not an unwarranted assumption, to be
sure.
But it is not essential, either.
Interesting in this regard are the slime molds native to Sol
III.444 At one stage in their life history, they are small and insignificant
one-celled flagellates capable of individual multiplication by simple fission.
At a later time, however, large clumps of these cells fuse together. The resulting
single, large, amoeba-like organism is capable of collective movement as a single
entity. During this "plasmodium" stage cell walls dissolve away, leaving
an amorphous mass of living protoplasm which can grow as large as 25 centimeters
or more.
Here’s the catch. Although there are many cell nuclei
floating around inside, the "body" of the slime mold in the plasmodium
stage bears no trace of the earlier division into many cells. The organism has,
quite literally, metamorphosed into a large single-celled organism.*
Hence, large extraterrestrial beings plausibly may be unicellular.
Another peculiar possibility is the diffuse-organism concept.
Despite the assertions by some that "no biological life has developed in
which a single integrated organism has covered large geographic areas,"600
it has been pointed out by the renowned American entomologist Edward O. Wilson
that a colony of social insects may be thought of as a kind of organism:
...weighing anywhere from less than one gram to as much
as a kilogram and possessing from about a hundred to a million or more tiny
mouths. It is an animal that forages amoeba-like over fixed territories a
few meters in extent.... The giant of all such superorganisms is a colony
of the African driver ant Dorylus wilverthi, which may contain as many as
22 million workers weighing a total of over twenty kilograms. Its columns
regularly patrol an area between 40,000 and 50,000 square meters in extent.565
What would an ET with detachable cells be like? Would we
recognize the parent organism as living? If it were intelligent, could it comprehend
us? Contact with an alien diffuse-organism might prove frustratingly difficult
for early human explorers of distant worlds.
[Note: See also the author's article "Extraterrestrial
Zoology", published in 1981.]
* This idea has already been used in several
science fiction stories by Larry Niven. His Bandersnatchi are an example of
an elephant-sized extraterrestrial organism constructed on a unicellular basis.
The creature consists entirely of "undifferentiated protoplasm."451
11.1
Specialization and Symmetry
Despite the fascinating possibilities suggested above, a compact
multicellular condition will probably develop on many worlds with highly organized
life.* This is primarily because multicellularity permits specialization.
Specialization permits a division of labor in the primitive
cell colony. Some cells become expert at, say, moving the body around. Others,
free to ignore motion, may perfect their skill at digesting foodstuffs. The
division of labor allows cells to do what each does best, and the aggregate
efficiency is vastly increased.
It is the business of life to accumulate information and complexity.
The specialized, highly differentiated organism is the bearer of far more information
than a more homogeneous or generalized entity. The specialist metazoan has greater
complexity.
This is not to say that complexity per se necessarily has
survival value. But solutions to complicated environmental crises may require
complicated solutions. As Carl Sagan observes, a solution involving many molecules
will frequently be qualitatively superior to one which involves only a few.20
There does seem to exist a general evolutionary trend towards increased information
storage, greater complexity, and higher levels of biological specialization.
By "specialization" we usually mean "the utilization
of specific structures which perform given functions within the body."
These structures are called organs.
Organs, while specialized in function, need not be specialized
in structure. It is entirely possible that extraterrestrial lifeforms might
have what is often referred to as "distributed organs." Instead of
neat packages of similar cells, distributed organs would be scattered throughout
the body. There are precedents for this on our own world.
For instance, insects inhale oxygen like other animals and
yet they have no lungs. Unlike the localized organs found in humans, mammals
and other animals, the insect "lung" permeates the entire body. The
insectile structure is crisscrossed, like a piece of Swiss cheese, with a network
of tracheal tubules and microscopic holes. Air reaches the interior cells only
by diffusion. The insects’ "lung" is a distributed, nonlocalized
"organ."
Peter Ritner has suggested that a creature’s brain might
be of a distributed nature.1550 We can imagine an enormous blob of
alien protoplasm suffused with more or less generalized nervous tissue. The
organism’s brain and body, like the breathing apparatus of the insect,
would be coextensive.
The number of organs of each type is highly variable. The
squid, for instance, has two kinds of hearts -- one for arterial blood and one
for the venous. Many terrestrial creatures have two or more eyes, multiple anuses
and stomachs, pairs of arms and legs, and so forth.
Sometimes, organs combine several functions at once -- such
as the human mouth. There is no reason to believe that ETs will have exactly
the same combinations as us.
They may have separate organs for eating, drinking, breathing
and speaking. The dolphin, for example, eats through its mouth, breathes through
its blowhole, and "speaks" through its "ears." The land
snail’s lung opens onto a passageway quite distinct from its food canal,
and sea cucumbers breathe through their rectums. (This is called "anal
respiration.") Brachiopods can only vomit their solid and liquid wastes,
because they have a "blind intestine," a kind of alimentary cul-de-sac.
Then there is the ant lion, which is incapable of voiding
any excrement at all. This is because all digestion occurs outside of its body,
by the powerful saliva it injects into its victims. After the prey’s innards
have turned to soup, the ant lion sucks the victim’s body clean of the
predigested pap, leaving a dry husk behind. There are no wastes.
And the members of phylum Nematomorpha (long worms)
have no mouths at all. All nutrients are "eaten" by direct absorption
through the skin. Aliens can be no less strange in their organ arrangements.
There is also the possibility of multiple-brained extraterrestrial
animals. After all, Brontosaurus and Diplodocus, two species
of giant dinosaurs, carried an enlarged section of neural tissue in the hip
region -- larger than the brain itself! The volume of this "sacral enlargement"
in Stegosaurus, another fossil animal of grand proportions, was perhaps
twenty times larger than the brain in the cranial cavity in the head. Further,
Homo sapiens (a species not yet extinct) tenders a curious "split
brain." The left and right hemispheres are two coordinated organs with
distinct functions and reactions to stimuli.502 Most vertebrate brains
have a similar dual construction.
In addition to varying degrees of specialization and organ
distribution, most extraterrestrial multicellular lifeforms will display some
sort of symmetry. Symmetry refers to the basic geometrical layout of an organism,
the way its various parts are positioned with respect to one another, its shape.
Probably the first to appear in evolution was radial symmetry.
The radiolarians -- small, spherical lifeforms -- are ideal examples. The main
body is distributed around a single point in a radiating fashion.
The cylinder shape is another form of radial symmetry, in
which parts are arrayed around a line instead of a point. Many sponges and coelenterates
display this pattern.
However, virtually all of the metazoans may be regarded as
having bilateral symmetry. In this design, an imaginary line drawn from head
to tail bisects the animal into two similar pieces. That is, the organism may
be divided down the middle by a plane which results in two mirror-image halves.
Humans are organized in this fashion, as are all mammals and most chordates.
Bilaterality, if we can trust the fossil record of a single specimen planet,
must have great selective value because it has been re-invented countless times
in most phyla of the animal kingdom.
Of course, body symmetry reflects lifestyle. Primitive sedentary
or attached animals are usually radial, a body configuration that permits exploration
of the environment in all directions. Conversely, animals that must pursue their
food more actively need a more mobile, streamlined shape. The bilateral cylinder
is ideal for efficient unidirectional movement through viscous media such as
water. And it is characteristic of the most successful of Earth’s fauna,
including all of the vertebrates and most of the invertebrates.
Based on the evidence of literally millions of terrestrial
animal species, we conclude that highly-organized mobile aliens will most probably
have bilateral symmetry if they have any physical form at all.
* Terrestrial organisms display remarkably
little variation in the sizes of the individual cells, most falling within a
single order of magnitude of scale. However, colder planets may spawn lifeforms
with somewhat smaller cells than ours. This is because there is less thermal
disruption at the molecular level at lower temperatures.63 But how small can
cellular building blocks be? The 1000 Angstrom "pleuropneumonia-like organism,"
or PPLO (the smallest living thing on planet Earth capable of independent metabolic
activity), is probably extremely close to the theoretical limit of roughly 400
Angstroms.1440,2395 An alien analogue to a human being, if constructed
of cells this small, would weigh a mere fifty milligrams and would stand only
five millimeters tall.
11.2
Xenobiomechanics
Besides the gross form alien life may assume, xenologists
also want to know how large these beings might be. What role does planetary
surface gravity play in determining the size and shape of bodies? What are the
limits of biological building materials? Will alien skeletal support structures
resemble our own?
As with most areas in the field of xenology, these questions
are quite complicated and have no quick, simple answers. A good place to begin,
however, is the Square-Cube Law, first recognized by Galileo more than three
centuries ago.2410
This mathematical law, which will hold anywhere in the universe,
states that volume increases faster than surface area as size is increased.
If an animal’s size is doubled, its surface area rises by a factor of
four. Its volume, and therefore its weight, rises as 23 or eight.
From this simple result flow many momentous consequences.
If weight increases eight-fold, but the surface which supported
that load increases only fourfold, then the pressure that must be sustained
to support the body against gravity has actually doubled. Bones are asked to
carry proportionally twice as much stress as before. Consequently, as animals
get larger, their bones must become thicker, sturdier, and squatter to accommodate
the higher pressures.
All parts of an animal must be modified when there is an increase
in size. Muscle tissue, the strength of which is determined by cross-sectional
area, must double in relative thickness in response to a twofold increase in
size. Lungs, kidneys, intestines and other blood-filtered organs also function
according to surface area rather than body volume. The entire organism must
be redesigned as it becomes larger.
The horror movies about giant insects ravaging the countryside
are really quite impossible. A bug the size of a house has a billion times more
mass to carry around, and its thin, spindly legs are called upon to sustain
pressures thousands of times greater than before. To walk at all, the misshapen
insect needs muscles proportionately thousands of times thicker than before
-- yet such tissue already virtually fills the hollow bones of the tiny original
so there is really no room to grow. If it did not collapse under its own weight,
or was not immobilized by the feebleness of its muscles, it would starve to
death because its stomach was a thousand-fold too small or would suffocate because
its tracheae could carry only a thousandth of the needed oxygen.
The worlds of size are truly worlds apart.*
* Consider the following excerpt from J. B.
S. Haldane’s Possible Worlds (1928):
A man coming out of the bath carries with him a film of water
of about one pound. A wet mouse has to carry about its own weight of water.
A wet fly has to lift many times its own weight, and a fly once wetted by
water or any other liquid is in a very serious position indeed. An insect
going for a drink is in as great danger as a man leaning out over a precipice
in search of food. If it once falls into the grip of the surface tension of
the water -- that is to say, gets wet -- it is likely to remain so until it drowns.
The majority keep well away from their drink by means of a long proboscis.974
11.2.1
The Challenge of Gravity
The strength of biological building materials sets an upper
limit on size. Galileo once calculated that a tree taller than about 100 meters
(on one-gee Earth) must buckle under its own weight. This is because its cross-sectional
strength would be insufficient to stabilize such a tall mass against collapse.
(Some sequoias, which slightly exceed this theoretical limit, are nevertheless
close to the maximum height attainable using woody materials.)
The typical loads sustainable by animal bone are about: 600
atm laterally (shear strength), 1000 atm in longitudinal tension, and as much
as 1700 atm in compression.204,1730 These should be compared to the
following: 340 atm for Douglas fir, 540 atm for hard-burned brick, and 5400
atm for cold-rolled steel.48,924 Clearly, bone is an excellent building
material.
The villain of this story is, of course, gravity. This force
at the surface of any world determines to some extent the maximum mass and size
of the animal life.
One way to meet this challenge is to take to the water. In
the sea, the force of gravity is partly cancelled out by the opposing natural
buoyancy of immersed bodies. There are no obstacles to large structures per
se.
But subtle problems arise when sea creatures grow too large.
It has been suggested that inertial mass, rather than weight mass, may be the
limiting factor. That is, the larger a body in motion is, the more it wants
to remain in motion. An extraterrestrial leviathan larger than a whale would
experience severe steering, braking, and turning difficulties. Cornering too
fast could easily exceed the strength of the building materials, and snap the
behemoth in two.400
There are also ecological considerations. Larger aliens will
generally eat more than smaller ones, all else being equal. Yet at the same
time it is getting huger, the organism is also getting bulkier and less maneuverable.
The animal needs to spend more and more of its day feeding. Indeed, the largest
whales must drive incessantly through food-laden waters in order to meet the
severe metabolic demands of their ponderous bodies.
Further, note that the mass which must be fed increases as
the cube of the linear dimension, the nutrients must be absorbed through the
surface area of a gut which increases only as the square of the linear dimension.
The Square-Cube Law thus predicts that at the same time it is becoming harder
to ingest food fast enough, it’s also becoming harder to actually utilize
what is eaten.*
What is the largest skeletonless creature that can exist?
We really don’t know, but in the ocean the answer is -- reasonably large.
Life without a rigid frame offers advantages difficult for
humans to fully appreciate. Many molluscs such as the squid and octopus have
pretty much lost their ancient shells and have become essentially skeletonless
lifeforms. Octopuses can stretch themselves quite thin, passing rubberlike through
small holes and narrow crevasses. Arms, eyes, and even head can alter shape
and elongate when necessary. The octopus has been called "the supreme escape
artist," and is known to be able to walk across desktops and the decks
of ships.
But a creature of the land must be a creature of gravity.
Whether resting on the surface or traveling across it, alien organisms must
find some means of support or be reduced to a groveling mass on the ground.
As long ago as 1917, the well-known zoologist D’Arcy
Wentworth Thompson speculated on the effects on evolution of altering the planetary
gravity. "Were the force of gravity to be doubled," Thompson declared,
...our bipedal form would be a failure, and the majority
of terrestrial animals would resemble short-legged saurians, or else serpents.
Birds and insects would suffer likewise, though with some compensation in
the increased density of the air. On the other hand, if gravity were halved,
we should get a lighter, slenderer, more active type, needing less energy,
less heat, less heart, less lungs, less blood. Gravity not only controls the
actions but also influences the forms of all save the least of organisms.958
The concept of the small, squat, muscular high-gravity beasts
and the tall, wiry, frail low-gravity beasts has been tediously reiterated by
generations of writers.
There are good reasons to doubt such simple conclusions.
First, we may properly assume that the strengths and densities
of biological building materials are roughly the same on any terrestrial planet
in our Galaxy. And it is certainly true that the maximum mass of a living organism
cannot exceed the crushing strength of its bones. But it is very important to
bear in mind that this observation is applicable only to maximum size. Indeed,
few Earthly animals exist at or even near the theoretical maximum.
This is because animals, extraterrestrial or otherwise, are
designed for motion. They must be able to withstand the peak pressures and accelerations
encountered during running and jumping. Standing at rest, for example, a horse
seems greatly overbuilt. But on the racetrack, where it may pull to a halt in
as little as 0.3 second (near the breaking point of its bones), its design limits
are more fully exploited.400
Second, it is relatively straightforward mathematically to
demonstrate, from the simple laws of Newtonian mechanics, that the maximum height
of animals on a planet is inversely proportional to gravity. That is, height
~ 1/g.214,1309 Similarly, we can show that the cross-sectional area
of supportive bone must increase directly with ~g, the bone radius as ~g1/2,
and the maximum mass as ~1/g3.
Let us consider the significance of these results.
We recall from an earlier
chapter
that the most massive of all terrestrial worlds should not have a surface gravity
in excess of 2.2 Earth-gees. (Although self-heating starless planets or superjovians
could have tremendous forces, even monstrous Jupiter only musters 2.64 gees.)
Luna, whose gravity is too feeble to hold sufficient oceans or atmosphere for
life, checks in at 0.16 gees. Apparently the range of plausible terrestrial
habitats for life spans a single order of magnitude of gravitational force.
With this in mind, let’s look at the maximum size of
land animals here on Earth -- a typically exotic, standard one-gee planet.
The largest land creature alive today is the African elephant,
weighing in at an impressive 6600 kg. But larger animals have trod the soil
of our world. The Baluchitherium, an extinct land mammal, had a total
mass of well over 12,000 kg. Tyrannosaurus rex, the largest land carnivore,
was about 13,500 kg.2409 The largest land animal ever was the Brachiosaurus,
weighing an estimated 45,000-78,000 kg, but we’ll ignore this majestic
brute because it is believed that he had to spend a great deal of time sitting
in swamps resting his tired bulk.
We shall hazard a crude guess, from these data alone, that
the heaviest viable exclusively land-dwelling creature that can plausibly be
designed on an average one-gee world is about 20,000 kg. (The precise value
selected doesn’t affect the conclusions very much.)
Applying the aforementioned maximum mass ~ 1/g3
relation, we discover the following: The weightiest alien animal that can inhabit
a 2.2-gee world might be about 1900 kg. On a tiny 0.16-gee world like Luna,
the largest creature could be nearly five million kilograms (though I’d
hate to try to keep it fed).
So on the heaviest of all reasonable terrestrial worlds,
animals such as walruses, small elephants, and even 70 kg humanoids are not
excluded. On massive, 2.2-gee planets, all animals the size of hippos or smaller
will certainly be possible with a modicum of redesign -- no need to call for
"powerfully built, squat creatures, perhaps rather like an armoured pancake
on multiple legs... limited to slow, creeping motions across the surface."41,45
There is no reason why such relatively minor alterations in surface gravity
should drastically affect the allowable sizes of typical alien animals.
This is not to suggest that gravity won’t control the
construction of large ETs to some extent. It is true that, in any given mass
category, the members of an animal species evolving on a high-gee world will
have shorter, stockier bones than those evolving in low-gee environments. We
can estimate how large this effect might be.
Consider the form of man. A typical human femur -- the most
perfectly cylindrical and the largest single bone in our bodies (found in the
thigh) -- is perhaps 3.5 cm in diameter. From the bone radius ~ g1/2
relation noted above, we find that the femur should increase to 5.2 cm on a
2.2-gee world, or fall to 1.4 cm on a 0.16-gee world, to provide equivalent
support for a 70 kg human body mass.
Such changes would probably necessitate major alterations
in bone distribution, structural stress loading, and internal organ design.
Experiments have shown that animals reared in high gravity environments tend
to grow slightly thicker than normal bones, stronger hearts, and to lose fat.
(See especially Kelly et al.,1309 Oyama and Platt,2418
Smith and Kelly,2419 Steel,2417 and Wunder.2416)
But by and large, alien creatures should not appear grossly
over- or under-built as compared with Earthly lifeforms of comparable mass.
* There is another still more subtle twist
to the story of pelagic lifeforms. On a high pressure world (which may or may
not correlate with higher gravity), more gaseous oxidant (e.g., oxygen) will
dissolve in the water in the oceans. The amount of O2 dissolved at
any given temperature is directly proportional to the atmospheric surface pressure
(Henry’s Law). So on a world with a hundred times the Earthly partial
pressure of oxygen, a hundred times more can dissolve in the sea. Warm-blooded
fish should be common. Cold-blooded fish without gills could also exist, breathing
directly through their skin like the earth worm and the salamander of Earth.
On a cold oceanic planet these effects would be even more pronounced, since
oxygen dissolves more readily in cold water than in hot.86,2513 And,
of course, more oxygen means larger bodies.
11.2.2
Meeting the Challenge: Skeletons
There are two most common varieties of skeleton currently
in use on planet Earth, both of which are suitable designs for extraterrestrial
land-dwellers.
The first of these is the exoskeleton, essentially a hollow
tube into which the creature’s viscera are poured. Our earnest ecological
competitors, the insects, have this form of body support.
The second type of skeletal structure, displayed by all vertebrates,
is called an endoskeleton. In this design, the support lies under the skin deep
within the body. The animal’s vital organs are hung around the central
spine like coats on a hat rack.
The two are complementary. Exoskeletons consist of gut surrounded
by skeleton; endoskeletons consist of skeleton surrounded by gut. Each is the
other turned inside out.
Which is the superior design for aliens?
It has often been pointed out by zoologists that a tubular
column of bone always gives greater strength than a solid beam of equivalent
mass.215,965 In virtually all situations involving static loading,
exoskeletons appear more advantageous (though usually only slightly).
For instance, the same resistance to bending is achieved with
an exoskeleton weighing half the mass of an endoskeleton.1715 Also,
the same mass of exoskeleton has seven times more resistance to buckling than
an endoskeleton -- although this advantage diminishes with size.1730
Finally, the area of bone to which muscles may be attached is far greater in
exoskeletons -- muscles may be better placed to take maximum advantage of the
mechanical advantages available in a given design.
Why, then, would any self-respecting extraterrestrial want
to be a vertebrate? If external skeletons are so clearly superior to internal
ones, why bother with the latter at all?
The answer is this: We’ve only considered static conditions.
Animals are designed not for the static situation but rather for the dynamic
one. As bulk size increases, endoskeletons soon outstrip exoskeletons because
their performance is superior under dynamic impact loading.1715,1730
Insects, the Earthly land-dwellers who make the greatest use of external skeletons,
are the same animals for whom gravity is least important. Small animals have
much less to fear from falling than do large animals.
That is, small animals, not needing high impact strength,
may develop the structurally sounder but dynamically less secure exoskeletal
arrangement. Large animals, whose greater size demands better protection from
destructive impact loads (e.g. falling out of trees), cannot afford this luxury.
The largest of all animals, both living and extinct, have had endoskeletons.
This is probably not a bad rule of thumb for alien lifeforms.
Of course, on a low-gee planet falling impacts would be somewhat
diminished for larger creatures. The exoskeleton might remain the preferred
design up to bigger sizes than we find on Earth.
The largest exoskeletons this world has ever seen measure
on the order of 10 cm (on land). It is not inconceivable that the most lightweight
of habitable planets could allow meter-sized or larger structures.*
There are, however, certain other disabilities of the exoskeleton.
Setting aside the usual extreme overmassiveness of the structure, the central
difficulty is the problem of growth. If an alien organism possesses a hard,
unyielding integument, there is no space left for its body to grow.
Insects on Earth solve this problem, albeit rather clumsily,
by periodically "molting" -- shedding the too-small, aging dermal
skeleton and replacing it with a newer, larger one. This procedure has the unhappy
consequence of leaving the animal highly vulnerable and less mobile during the
time between the actual molt and the completion of the replacement. Predators
lick their chops.
ETs may have found better solutions. For instance, an exoskeleton
could be constructed from a series of overlapping laminar plates. They would
be designed in adjustable sections held fast by a protein glue. This glue could
then be loosened from time to time to permit the plates to slide a bit farther
apart, thus enlarging the interior volume of the skeletal cavity and permitting
continued growth.
Another alternative for aliens might be to use a hard, rubberlike
substance in the outer dermal layers. This material could be forced slowly to
expand (by biochemical means) while providing continuing firm support. Similarly,
one can imagine specialized polymers with different unidirectional cohesion.
Maximum tensile strength would lie in the horizontal plane of the ET, and minimum
strength would lie in the vertical direction -- thus permitting vertical growth.
Extraterrestrials may have an exoskeleton which, like the
human skull, has a very thin layer of living tissue over it. As the organism
grew, material would be dissolved from the interior face of the skeletal wall
and redeposited as fresh bone on the outward growing side. Massiveness would
represent an increasingly serious problem with size, but any significant thinning
of this "shell skeleton" would render the animal more prone to lethal
puncture.
Higher lifeforms elsewhere in the Galaxy may not be limited
to only two choices -- exoskeleton or endoskeleton. Countless outré support
structures may be readily imagined, but it will suffice to mention a mere handful
of them here.
One of the most popular alternatives, which appears in the
drawing on the following page, is known as the "basket skeleton."
(Figure 11.1) Found only in the echinoderms of Earth -- sea cucumbers, starfishes
and sea urchins -- have a bone structure which is neither endo nor exo but rather
a peculiar combination of both.
Figure 11.1 Examples of the Basket
Skeleton
|

|
Metacarpal endoskeletal bone from the wing of a
vulture.958 This internal structure is stiffened after the
manner of a Warren’s truss in mechanical engineering, providing
both strength and lightness.
A possible large-animal analogue of the basket
skeleton? |
|
Scanning electron micrograph of the basket skeleton
structures in the sea urchin Echinus esculentus.215 In
life, the interstitial spaces are filled with living cells which keep
the bony surfaces smooth. |

|
|
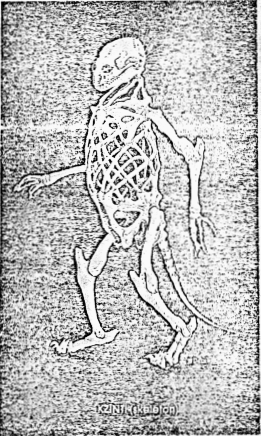
|
THE BASKET SKELETON
(Dalzell and Niven2421)
|
Instead of a central support column or an exterior support
tube, the basket skeleton is like a piece of calcified Swiss cheese, a kind
of bony trellis. This frame is a curving, intricate labyrinth of molded biocrystal.2191
Each segment of the internal basketwork is strapped to the rest with sturdy
collagen fibers looping through the structure "like laces through eyelets."215
The idea of intelligent aliens with basket bones appeared
in science fiction as long ago as 1937, in a novel by Olaf Stapledon:
Then there were men that had developed from a slug-like
ancestor along a line which was not vertebrate, still less mammalian. Men
of this type attained the necessary rigidity and flexibility of limb by means
of a delicate internal "basketwork" of wiry bones...1946
Another alternative is the multiple endoskeleton. Instead
of one spinal column, aliens may sport two or more major internal support columns.
On Earth the flatworm and other free-living turbellarians
have a double spinal cord which runs the length of their bodies. While these
twin structures serve a neural rather than a support function, the evolutionary
implications are clear: ETs may have at least two, possible more, spines.
Such a "ladder skeleton" would provide added postural
stability, strength, and reliability. Although turning or twisting motions of
the trunk might be somewhat restricted (even if the multiple support posts were
segmented or jointed), much heavier loads could be hefted by this physically
powerful creature. Ladder skeletons may actually be selectively advantageous
in many niches on high-gravity worlds.
A still different arrangement is another analogy to terrestrial
experience. The "hydrostatic skeleton," as it is customarily called,
is typically found in rather small marine organisms such as earthworms and nematodes.
(Internal pressurization is also found in echinoderms, some molluscs, caterpillars
and spiders, although to a lesser extent.)
Instead of bone or cartilage, the hydrostatic animal is supported
by pressurized fluid in the interior. The idea is to make use of the incompressibility
of water. Any closed container with flexible walls will serve; such a body is
really just a bag of water which can change its shape but not its volume.
Although perhaps restricted to smallish lifeforms on other
worlds, the hydrostatic skeleton might possibly be scaled up dramatically if
a more viscous support fluid than water could be found. This liquid skeleton
would be held in place inside a tough fiber-strengthened central tube with extensive
reinforcing musculature. And the possibility of devising muscles that push instead
of pull has also been raised.878
Then there is the "corkscrew skeleton," typified
on this planet by several of the nemertines. These are predominantly marine,
bottom-dwelling wormlike organisms typically 20 cm in length (a few have measured
several meters in length when fully extended). Their bodies are soft and extensible,
with no exoskeleton or vertebral structures whatsoever.
Instead, these creatures have evolved a helical arrangement
of firm collagen fibers just beneath the upper layers of the skin. This spiral
skeletal construction, although relatively weak, allows large changes in shape
in response to the contraction of circular and longitudinal muscles by simply
changing the pitch of the collagen helices.1730
It is not difficult to imagine scaling up the corkscrew skeleton
to larger sizes for serpentine creatures on other worlds. The tremendous stabilizing
effects of internal tissue make this possible. Vertebrate tendon has a tensile
strength approaching 1000 atm, which compares favorably with bone and insect
chitin. Further, it is well-known that muscle tensions are frequently much greater
than the loads imposed on the bones.215 The extensive interior sinewy
bracing that would be required for adequate support is made more plausible by
the observation that the human body itself has more than 400 individual muscles,
representing roughly 40% of the total body weight.
Countless other skeletal schemes may be visualized, including
such oddities as telescoping bones (imagine a giraffe with a retractable neck),
limbs with universal joints (a full 360º freedom of motion, like an owl’s
neck), and variable skeletons (an ability to restructure and reallocate the
internal distribution of bones). Designing ETs is an enjoyable game for anyone
with a sharp imagination, and can prove both fun and informative if you stick
to the rules.
* These limitations are far less severe in
the sea. Fossil marine arthropods have been measured up to three meters in length
-- most of it hard exoskeleton. Twenty-meter-long invertebrate molluscan squids
(e.g., Architeuthis princeps) likewise exist with large external carapaces,
although most of this great size consists of soft, fleshy tentacles rather than
hard, rigid exoskeleton.
11.3
Alien Locomotion
Mobility seems to be a central characteristic of all large
animal lifeforms on Earth. It may be presumed that this function will be evolved
elsewhere in our Galaxy in delightfully diverse ways.
There are three distinct media which may be used for locomotion
in any planetary environment: Sea, land, and air. Life is believed to have arisen
in the oceans, and is likely to have done so again on uncounted worlds among
the stars. The first creatures that ever moved evolved in the seas, so it is
appropriate that we begin with aquatic locomotion.
The intention here is merely to illustrate, not to exhaust,
the possibilities.
11.3.1
Aquatic Locomotion
The most ancient and respected form of aquatic locomotion
is by undulatory movements of the body. Even at the level of the protozoans
we find this to be true. Alien fishes in watery oceans of other worlds must
often have evolved some similar technique.
Earth’s first fish species -- eels, lampreys, and so
forth -- swam in a serpentine fashion using sinuous motions of the body. More
evolved creatures of the sea, such as the stingray and the skates, drive forward
through the medium by rippling their bodies in a series of back-moving up-and-down
waves (rather than the sideways motions of the eel).
As time went on, still more sophisticated methods came into
general usage. Aquatic lifeforms developed paired fins for added power and control.
Stabilizing vanes appeared. As in the case of the alligator and several other
water-dwelling lizards, many fish use their tails alone as the main source of
thrust.*
When traveling at a steady pace, the output of propulsive
energy is proportional to the resistance of the surrounding medium. Therefore,
it is in the best interests of the seagoing organism to reduce that resistance
as much as possible. How might alien lifeforms accomplish this?
We might take a tip from the dolphins. These lissome marine
mammals have a unique conformable skin which changes shape slightly as the animal
slices through the water. At higher speeds, the cetacean varies its skin surface
to maintain an exactly hydrodynamic streamlined form. The smooth, laminar flow
of water over the dolphin’s body minimizes resistance and saves large
amounts of energy. Indeed, porpoises are known capable of steady speeds of 55
kph, and still higher velocities are probably attainable by schooling.1708
But there is no need to hurry so. Most animals manage quite
well with far less speed at their command. Earthworms, nematodes and others
with hydrostatic skeletons slither along the ocean bottom like tiny accordions.
Jellyfish, squids, cuttlefish and octopuses all use a kind
of biological ramjet for propulsion. After water is passed over the gills for
respiratory purposes, the exhaust is rapidly expelled. The siphon-like apparatus
is such that, by merely gasping harder and faster, the organism can propel itself
backwards in a series of sharp bursts. Octopuses can manage 8 kph or better
in this fashion, and there is no reason why extraterrestrial biological ramjets
could not do far better than this.
There are more exotic possibilities. One highly unusual technique
which has never been exploited on Earth is osmotic power.
A permeable sac filled with salty water and placed in a beaker
of fresh water will expand. This is the process of osmosis at work: The pure
solvent flows through the membrane into the region of higher salt content. This
represents a force which, when distributed over the surface of the sac, becomes
a pressure.
Pressure is a force which can be harnessed to do useful work.
And osmotic pressures are usually quite high: The pure water in the example
above tries to dilute the seawater with a pressure of nearly 25 atm.230
This osmotic force is known to increase directly both with temperature and with
salt concentration in the sac.
Could osmosis drive an alien fish through some faraway ocean?
Imagine a "freshwater" ET constructed in three
sections: Head, torso, and hindbulb. The head faces forward, exposing a sac
of highly concentrated salts to the surrounding water. An osmotic pressure of
many tens of atmospheres forces the pure liquid into the sac in an attempt to
dilute the salts. But this working fluid is continuously filtered through a
large organ in the torso. There, energy is expended to reconcentrate the saltwater
and to extract the excess water, which is stored in the hindbulb. When the creature
wants to move, this liquid is rapidly exhausted from the hindbulb through a
small nozzle in the tail.
There are three evolutionary preconditions for the emergence
of the osmofish.
First, there must be some reason why the animal cannot simply
inhale the surrounding fluids directly and jet them out again. Perhaps the osmofish
inhabits a sea filled with poisons, or maybe the creature’s organs might
be thermally or ionically damaged if contact with the outside were permitted.
There is some terrestrial precedent for this: To this day, cell nuclei exposed
to life-giving oxygen are poisoned by it.
Second, the medium must be so viscous that ordinary fin-flapping
and body-wriggling are wholly inadequate. On a sulfur thalassogen world, where
the sea alternated regularly from very thick to very thin, osmotic propulsion
might evolve as a backup system for when the oceans became too gluey to swim
in.
Third, the diffusion process across the membrane must be
sufficiently fast to render the osmotic drive competitive with other forms of
locomotion. This is probably do-able.
The osmofish is thus a very real and plausible possibility.
The surfaces of seas and inland pools of other worlds may
harbor still more surprises. There are many forces that may be tapped for motive
power. Let us consider just a few of these.
The swamps of Earth are inhabited by a fascinating variety
of water beetle known to zoologists as Stenus. Thrown to the middle
of a pond, these tiny creatures shoot to the safety of the banks at the water’s
edge. The method of propulsion is not unlike that used by toy camphor boats:
surface tension.
Surface tension is a property of all liquids, causing them
to adhere to themselves at the interface between fluid and atmosphere. This
tension represents a considerable force, and many insects such as water striders
are elevated above water entirely by this support.
Stenus not only uses the surface for support, but
for propulsion as well. By secreting a substance similar in action to camphor,
the surface tension behind the organism is lowered. The resulting imbalance
of forces causes the beetle to be drawn forward rapidly.
The data in Table 11.1 suggest that few liquids can compete
with water in terms of providing useful energy for surface tension locomotion.
Picture a propulsive appendage with reduced tension on one side only. Table
11.1 gives the maximum available force along each meter of appendage for which
the surface tension is lowered from its normal value down to zero.
|
Table 11.1 Surface Tension and Alien Locomotion
|
|
Liquid |
Typical Surface Tension |
Liquid |
Typical Surface Tension |
| |
(Newtons/meter)
|
|
(Newtons/meter)
|
|
Iron, molten |
1.60
|
Hydrogen iodide |
0.028
|
|
Mercury, liquid |
0.450
|
Hydrogen bromide |
0.027
|
|
Lead, molten |
0.400
|
Chloroform |
0.027
|
|
Selenium, molten |
0.092
|
Carbon tetrachloride |
0.026
|
|
Hydrazine |
0.092
|
Methylamine |
0.025
|
|
Sulfur, molten (max.) |
0.073
|
Hydrogen chloride |
0.024
|
|
WATER |
0.073
|
Ethanol |
0.024
|
|
Glycerol |
0.060
|
Methanol |
0.022
|
|
Formamide |
0.058
|
Ammonia |
0.020
|
|
Sulfuric acid |
0.055
|
Hydrogen cyanide |
0.018
|
|
Bromine |
0.040
|
Fluorine |
0.016
|
|
Formic acid |
0.038
|
Methyl chloride |
0.016
|
|
Iodine |
0.035
|
Oxygen |
0.013
|
|
Sulfur dioxide |
0.033
|
Argon |
0.013
|
|
Carbon disulfide |
0.032
|
Carbon monoxide |
0.010
|
|
Acetic acid |
0.030
|
Carbon dioxide |
0.001-0.01
|
|
Chlorine |
0.028
|
Neon |
0.0055
|
|
Nitrogen tetroxide |
0.028
|
Helium (4.15°K) |
0.00012
|
How large could an alien surface-sprinter be?
Let’s assume 100% efficiency, propulsive appendages
one millimeter in radius with the approximate density of water, and a total
length of all propulsive strands of a hundred meters when unfurled.
If such a creature totaled 2 kg in mass, it would theoretically
be capable of accelerative bursts on the order of 0.4 gees -- the approximate
rate achieved by Olympic-class human sprinters. The bundled threadlike appendages
would represent 16% of the total body mass in this case. If only a tenth as
much acceleration is required, our surface tension beast may weigh as much as
20 kg, with appendages a mere 1.6% of the total.
There are other forms of surface locomotion. The basilisk
lizard of Central America has the unique ability literally to walk on water.
(Locally, this has earned it the appellation "lagarto Jesus Cristo.")
This small but active reptile, biologically related to the
desert iguana, speeds across lakes with a gait suggestive of the hasty canter
of a terrestrial biped. This ability is of tremendous selective value, since
by scurrying along the water’s surface the animal avoids aquatic predators
as well as its enemies back on shore.2433
The basilisk is a cold-blooded animal, so a warm-blooded
ET might be expected to do much better. The coordination and agility required
at such high speeds could provide the challenge necessary for the evolution
of intelligence on another world.
What about the principle of the rowboat? Rowing consists
of a series of power strokes by which oars move backwards relative to the boat,
thus driving it forward. Is it possible that extraterrestrial animals could
copy this idea and, fantastic as it may sound, row across the surface of the
sea?
Mother Nature has provided ample precedent. The hexapodal
water beetles Dysticus and Acilius, and an insect known as
the water boatman (Notonecta), use their middle and hind legs to row
themselves along near the water’s surface. Hairs on their appendages have
a distinctive oarlike appearance, and it has been calculated that this method
of locomotion could be as much as 45% efficient energetically.230
The platypus, a far larger animal, may also be said to "row."
It is often asserted that rotating structures such as flywheels
and paddlewheels cannot be used in living organisms, because "all parts
of the body must be connected by blood vessels and nerves." This simple
objection contains two hidden fallacies.
First, it is now known that certain spirochetes have flagellae
driven by tiny ionic motors complete with rotor, stator, bushings and drive
shafts.2432 The feisty bacterium E. coli, for instance,
comes equipped with a rotor spinning at roughly 60 cycles per second (just like
the alternating current from a wall socket). Although admittedly small in size,
this micromotor involves an axle which spins around its long axis through a
kind of universal joint. It is in no way connected directly to the main body
-- save through the rotating armature itself.1661
This finding contradicts the common assertion that living
organisms may not contain detached, self-rotating parts.
Second, there really is no need to design a paddlewheel as
a true rotating assembly. Fins may be stuck out, scooped through the water,
and retracted again in regular sequence. This would provide both the appearance
and the motive power of a true paddlewheel.
Many strong paddlers have evolved on this planet. The duck,
for example, is efficient enough to propel herself through the water at a stately
three kilometers per hour.1006
Paddlewheel aliens thus may not be excluded a priori.
Wind power is yet another largely unexplored avenue of water
surface locomotion. The possibilities are barely hinted at by the fauna of this
planet. Whales are known playfully to dive underwater, leaving only their giant
broadleaf tails exposed above the surface. These lighthearted leviathans then
"sail," catching gusts of air on their huge tail vanes and drifting
with the wind for hundreds of meters before they’re forced to come up
for air.5522 (Since it is far more efficient to swim than to sail,
this is presumably a form of play.)
Let’s carry the idea to its logical conclusion.1946
Imagine a mollusc-like organism with a moderately thick concave
shell which inhabits the coastal shallows of another world. Over the years this
alien species acquires the ability to float boatlike on the inverted shell,
drifting with the shore currents slowly across the face of the planet. Such
creatures might feed on floating plant life such as surface scum or the tops
of seaweed stalks.
With evolution, the shell might become better adapted for
navigation, perhaps developing a more streamlined underbelly. This would allow
the ET to better chart its course between patches of food. Eventually it would
discover that its speed could be significantly augmented with a crude "sail,"
a thin membrane growing up out of the animal’s back.
In time, the membrane could become retractable, or even delicately
manipulable by muscles. With the emergence of a brain and sensory organs strictly
comparable to molluscs on this planet, a kind of living clipper ship might evolve
-- complete with masthead (forward sensors), jib, mainsail and riggings (extensible
tendon), and probably a rudder. The minds of such lifeforms could be truly awesome.
What other glorious mysteries may await us in the star-dusted
depths of space?
* As a general rule of thumb, the top swimming
speed of aquatic lifeforms on Earth is roughly ten body-lengths/second. Normal
cruising speeds check in at about four body-lengths/second.224,2424
11.3.2
Travel by Land
The surface of a terrestrial planet is a very crude, rugged
environment, vastly less homogeneous than either sea or air. There are rivers
to be forded, forests to be traversed, craggy crevasses to be leaped, and sand
and mud and swampy bogs to be waded. Travel by land thus demands the development
of extremely versatile locomotive techniques.
Xenologists find Earthly zoology most instructive in this
regard. Here, the vertebrates and the arthropods are the only major animal groups
which have proved adaptable enough to fill virtually all available planetary
niches. Curiously, they are also the only two groups that have heavily exploited
the mechanical principle of rigid levers in locomotion. The implication seems
to be: Successful phyla use struts.
The ambulatory limb is probably the closest thing to a "universal"
locomotive device for surface travel. But how many legs will aliens have (Table
11.2)?
|
Figure 11.2 How Many Legs?
|
|
To be classified as in N-ped, an animal must either;
(1) Have N limbs used for walking or grasping,
or (2) customarily ambulate or stand on N limbs. Arguable
cases are enclosed in brackets
|
|
NULLIPEDS (0 legs) |
--
|
Snakes |
|
MONOPEDS (1 leg) |
--
|
Freshwater clam; snail; sea horse; rotifer |
|
BIPEDS (2 legs) |
--
|
Humans; kangaroos; seals; many small mammals (squirrels,
Arctic hares. etc.); birds (penguins, ostriches, most common Aves);
many extinct dinosaurs (notably Tyrannosaurs, Anatosaurs, Hadrosaurs,
Camptosaurs, and Astrodemus); sea cucumber larvae (Cucumaria
frondosa) |
|
TRIPEDS (3 legs) |
--
|
(Kangaroo); {Tyrannosaurus} |
|
QUADRUPEDS (4 legs) |
-- |
Most land-dwelling chordates (mammals, reptiles, and
amphibians); most primates; seals; Arctic hares; inchworm; (praying
mantis} |
|
PENTAPEDS (5 legs) |
--
|
Starfish; platyrrhine ‘prehensile-tail’
monkeys (Cebidae); howler monkeys (Alouatta); capuchin
monkeys (Cebus); woolly monkeys (Lagothrix); woolly spider
monkeys (Brachyteles); spider monkeys (Ateles); {elephants} |
|
HEXAPODS (6 legs) |
--
|
Many insects (cockroaches, flies, ants, chiggers, etc.);
larval myriapods (Pauropoda) |
|
HEPTAPODS (7 legs) |
--
|
Springtail "catapult insects" (Collembola); (Ichneumon
fly (with ovipositor)} |
|
OCTOPEDS (8 legs) |
--
|
Octopus; shrimps (ambulatory appendages only); tarantula;
scorpion (Vejovis); spiders; sea spiders (Pycnogonida);
spider mites; fowl ticks |
|
NONAPEDS (9 legs) |
--
|
{Scorpions} |
|
DECAPODS (10 legs) |
--
|
Squid; "Horseshoe" king crab (Xiphosura); spiny
lobster; scorpions (counting pincers but not tail); Carribean and Antarctic
sea spider species; geometer or looper caterpillars |
|
MULTIPEDES (many) |
--
|
Scorpions (all appendages), 11 "legs"; one species of
Antarctic sea spider (Pycnogonida), 12 legs; dipteran larvae
(Liponeura), 12 legs; caterpillars, 12 legs, 14 legs, but most
typically 16 legs; myriapod adults (Pauropoda), 18 legs; shrimps
(all appendages), 22 legs; tracheate myriapods (Symphyla), 24
legs; centipede (Lithobius forficatus), ~30 legs; "house centipede"
(Scutigera coleoptrata). ~30 legs; centipedes (Scolopendra
morsitans), ~40 legs; millipedes (Polydesmida), ~40 legs;
chambered nautilus, ~60-90 "legs"; extinct trilobite, ~100 legs; one
slow-moving centipede (Orya barbarica), ~200 legs; various millipede
species, ~64-400 legs |
On strictly mechanical grounds, three points are needed to
geometrically define a surface plane. Two points define only a line. Any ET
attempting to stand on only one or two points of contact must be unstable --
almost by definition. A minimum of three legs would seem to be necessary.
Tripedalism appears to have little to recommend it. All organisms
will be designed for dynamic rather than static conditions. But when the three
legger walks, it must lift one foot off the ground. The instant it does so,
it is no longer supported by a three point platform but merely by two -- which
is dynamically unstable. From an engineering standpoint, four legs would allow
the organism to remain balanced while one leg was moved to walk.
Still, the creature may not mind unstable walking. Bipedalism
has been quite common among the birds, reptiles, and mammals of Earth, and should
actually be favored on low-gee worlds.736 But it seems difficult
to plead for the existence of extraterrestrial tripeds when two legs seem easier
to operate and maintain.86 And then there is the old argument that
appendages will always come in pairs, because of our origins in the sea and
the need for hydrodynamic symmetry.2435,2436,2437
However, xenologists remain unconvinced by such reasoning.
They dismiss the stability problem as academic, recalling that most running
bipeds and quadrupeds keep only one or two limbs on the ground during the locomotary
cycle (as in the pace gait, or the trot, the rack, or the gallop). Also, terrestrial
animals in the same general weight class also have roughly equivalent vestibular
balancing equipment, so going from three to two legs probably wouldn’t
save much there.
As for the objections to unpaired limbs, they are pragmatic
but unpersuasive.* Many dinosaurs, such as Tyrannosaurus rex, and a
few large contemporary organisms, such as the kangaroo, run bipedally but stand
tripedally. These creatures’ tails are as thick and strong as the forelegs,
and are regularly used as postural support. Furthermore, when kangaroos fight
they are known to rear up on their tail to free both legs in order to deliver
crushing kicks at their opponents.450 In some sense, then, these massive marsupials
may be considered "facultative tripeds."
Pure tripedalism appears rather rare on Earth, but this is
insufficient to rule it out elsewhere. Advanced aliens on other planets may
have different evolutionary ancestors than we. For these reasons, xenologists
expect to find at least a few intelligent three-legged species in our Galaxy.
At the heart of the multipedia controversy is the issue of
neural control. How much independence of movement is each appendage to have?
Must quality be sacrificed for quantity?
Humans and other primates, basically quadruped in design,
have: full and complete control of each of the four limbs. But there is some
recent evidence that other four-leggers are not so fortunate.
At moderate speeds, horses appear to be naturally predisposed
toward either of two distinct natural gaits. One is the trot, in which the diagonal
legs swing in unison. The other is the pace, with the two limbs on the same
side swinging together. It turns out that a horse which trots cannot be made
to pace, and vice versa, without extensive training in special harnesses.400
So even among quadrupeds there is a hint of pre-programmed
leg motions. Many otherwise perfectly acceptable gaits are generally ignored
-- four-leggers have twelve symmetric gaits available, but only about four of
these are used much at all.2439 The hexapodal insects are still more
wasteful. With a total of some thirty-six symmetric gaits available, cockroaches
and flies use just two. Limb movements have become almost totally "hard-wired."
The extreme case of the centipede and millipede drives this
point home. These creatures perambulate by sending a single signal pulse the
length of their partitioned bodies. As the message reaches each segment in turn,
the connected limbs automatically sweep forward, robotlike, in a downsloping
arc. (Otherwise, the appendages do not move at all.) It is quite impossible
for a millipede to wiggle just one leg.
As a general rule applicable to extraterrestrial lifeforms,
then, we might suspect that fewer limbs means more control per limb. But why?
The late John Campbell and others have suggested that extra arms and legs means
extra demands on the brain. Six legs are impossible in large aliens, asserted
Campbell, because there would be substantial neural-coordination problems in
guiding so many limbs.1380 No brain could meet such an enormous challenge.
Most xenologists today would probably dispute this contention.400
The neurological equipment needed to operate an additional appendage is far
less complicated than the circuitry required for, say, an extra eye. While the
processing of visual data takes millions of neuron interconnections in the typical
mammal, that same organism requires only on the order of thousands to actuate
the muscles independently.501 We see that only a relatively small
slice of the brain is dedicated exclusively to motor control, whereas about
one-third of the entire organ is wholly committed to sensory functions. It should
therefore be orders of magnitude less difficult to add extra arms and legs to
ETs than extra eyes.
There are many who seriously believe that hexapodal aliens
are quite plausible.1216 One recent fan of hexapedia is Bonnie Dalzell,
a writer trained in paleontology who has been called "the best designer
of alien life in the United States."2423
Ms. Dalzell insists that vertebrates on Earth have four limbs
solely because of their common descent from fishes adapted to free-swimming
conditions in large, open oceans. Fish needed only two sets of independent diving
planes to maintain stability as their powerful tails drove them through the
water. Perhaps if we had evolved from the Euthacanthus, a fish which
lived in the Devonian Period with no fewer than seven pairs of fins, we might
be hexapodal or more-podal today ourselves.1222
So, assuming ETs have limbs, how many digits (i.e., fingers,
toes) are they likely to possess (Figure 11.3)?
|
Figure 11.3 How Many Fingers?
|
|
MONODACTYL (1 finger) |
--
|
Horses; Thoatherium (an extinct mammal the
size of a small dog),. (shrimps); (octopuses) |
|
DIDACTYL (2 fingers) |
--
|
Camels; sheep; pigs; deer; caribou; pronghorn cattle;
giraffe; goats; antelopes; ostriches (massive running bird); Choeropus;
Cyclothurus; some dinosaurs |
|
TRIDACTYL (3 fingers) |
--
|
Rhea and cassowary (massive running birds); bandicoots
(Peramelidae); many extinct mammals (Litopterna, Chalicotheres);
extinct horse (Paleotheres); extinct rhinos (Hydracodonts);
extinct reptiles (Struthiomimus); most dinosaurs (Archeosaurus,
etc.); rhinoceros |
|
TETRADACTYL (4 fingers)
|
--
|
Some primates (Colobus, Ateles); sloths; ancestral
horse (Eohippus); hyenas; Cape hunting dogs; tapirs; hippopotamus;
elephant shrews; octodont rodents; armadillos; chickens and most birds;
extinct mammals (Titanotheres); theropod saurischan dinosaurs |
|
PENTADACTYL. (5 fingers)
|
--
|
Humans; most carnivores; most reptiles; walrus; dolphin;
elephant; armadillo; sauropod dinosaurs |
|
MULTIDACTYL. (>5 fingers)
|
--
|
Many fish species, e.g., Sargassum
(10 fingers) |
It is generally believed by paleontologists that the original
amphibian, ancestor to virtually all lifeforms on land, had a hand with five
digits. However, it has been admitted that this really cannot be accurately
determined because of "the imperfect state of the earlier fossils."223
Many authors have postulated that one or two additional digits might have existed.
In fact, a few fossils are known in all three major tetrapod
classes -- mammals, reptiles, and amphibians -- which appear to have marginal
bony vestiges from a piscine ancestor.223 There is precedent for
multidactylism among the fishes. The Sargassum fish, to take one example
of many, possesses pectoral fins which operate like tiny ten-fingered (decadactyl)
hands.586 But this does seem to push close to the upper limit: It
is difficult to understand how more than ten digits could be of utility to any
animal.
There is a general evolutionary trend toward a reduction
in the number of digits, especially in runners and swimmers. The adaptation
to running commonly proceeds in two stages. Dactyls are lost when the animal
first switches to running, taking on a "digitigrade locomotion" --
walking high up on the fingers. In later stages, the animal progresses to "unguligrade
locomotion" -- or walking on the fingernails (hooves).
The size of hooves is markedly influenced by the environment.
Dwellers on shifting sands and mushy marshes develop larger, flatter ungulae.
Mountain dwellers, on the other hand, retain smaller, pointed, digit-like hooves.
For instance, the African kopjes appear to be standing on tiptoe -- a specialization
for rocky terrain and for taking advantage of every slight irregularity along
narrow ledges on cliffs.223 Ungulates tend to be herbivores.
The ancestral mammal is thought to have been pentadactylate
with an (at least partially) opposable thumb. Unfortunately, when dactyls are
lost this is always the first to go. But exactly how does an animal lose its
thumb? The first digit vanishes more often in the foot than in the hand, which
suggests that you will lose your thumb if you walk on it. Also, animals who
use their extremities for non-arboreal progression -- such as locomotion across
level ground rather than by swinging through the trees -- do not retain the
digit. However, the mandate to remain arboreal is not infallible. The thumb
has retrogressed in many arboreal forms (such as the sloth) when the digit is
not used for grasping. Reduction in number seems to be the penalty for disuse.223
Environment plays a key role in determining what evolves.
For instance, Dalzell expects to find six-leggers on worlds with small, shallow
oceans. There, bottom-dwelling fishes would be the predominant marine lifeforms
early in evolutionary history, occupying coastal and freshwater environments.
If the planet had a very seasonal climate, perhaps accompanied by large-scale
periodic evaporation of shallow seas, then few fishes would have the chance
to evolve into highly efficient swimmers -- as evidently occurred on Earth.
With little open ocean available during most of the year,
potentially adept swimmers might not find it a very profitable niche to occupy.
The bottom-dwelling many-finned fishes and crabs would inherit the land instead,
and go on to produce a rich variety of hexapodal alien lifeforms there.
Many advantages can be cited in favor of hexapedia. On a
high gravity world, for instance, each leg would support far less mechanical
stress than those of a quadruped of similar mass in the same environment. This
should be of great selective value. An additional consideration is that the
loss of one limb through accident or misadventure would be less serious for
six-leggers than four-leggers. It’s always good to have spares.
Another advantage is better balance. Hexapodal locomotion
provides a stable support tripod for the ET even at high speeds, unlike quadrupedal
running.2605 And as pointed out earlier, there should be no problems
with coordination, Says Dalzell: "Earthly insects with three pairs of legs
are hardly noted for their well-developed mental powers but most of them walk
just fine."736
Prophetically, Sir Richard Owen, a British paleontologist
of some repute, wrote nearly a century and a half ago (1849):
We have been accustomed to regard the vertebrate animals
as being characterized by the limitation of their limbs to two pairs, and
it is true that no more diverging appendages are developed for station, locomotion,
and manipulation. But the rudiments of many more pairs are present in many
species. And though they may never be developed as such on this planet, it
is quite conceivable that certain of them may be so developed, if the vertebrate
type should be that on which any of the inhabitants of other planets are organized.
The conceivable modifications of the vertebrate archetype are very far from
being exhausted by any of the forms that now inhabit the Earth, or that are
known to have existed here at any period.2422
Despite the tremendous versatility of legs, there are many
other kinds of locomotion possible on land in specialized niches. The freshwater
clam, for example, has a single hatchet-shaped muscular foot protruding from
its shell which allows it to plow along the bottom at a leisurely pace.
As the organism "walks" its foot is thrust forward
through the bivalve shell. Blood flows into many sinuses, causing the organ
to expand and anchor the animal securely. Retractor muscles contract, pulling
the clam a few centimeters forward. Blood is then drained from the foot, which
shrinks and is withdrawn from the sand. The cycle repeats.
The giant clam of Earth has reached masses of more than a
third of a ton. It is entirely possible that huge pelagic lifeforms may scour
the bottoms of alien lakes and streams in this fashion.
Another monoped is the common snail, which slides along on
a cushion of slime. Its movements are similar to those of the clam, but its
foot doesn’t appear to move much at all. This is because the snail takes
shorter "steps." The creature advances in a peculiar loping motion
by forcing a wave of tension down the length of its foot. This oscillatory thrusting
causes it to slide gently forward.
One of Larry Niven’s fictional extraterrestrials, the
Bandersnatchi, locomotes in this way.
Although there are no direct large-animal analogues of the
protozoan pseudopods of amoeboid movement,** the "tube feet" of the
starfish might be considered a distantly related metazoan form. Starfish, sliding
silently over rocky surfaces, give no hint of the source of the graceful fluidity
of motion. But upon turning one on its back, one sees hundreds of tiny sucker-tipped
tubes lining the underbelly. These are the tube feet, opening into a water-filled
canal running the length of each of the five arms.
By pumping water in or out of the canal, the organism can
control the actions of its feet. Motion is very rapid: The tubes are pushed
forward in the direction of travel and then fixed by their suckers to the rock.
Successive waves of tension and relaxation waft the animal along. ETs too may
try this trick.
There are many alternatives that have never been tried on
Earth. A world covered with vast tracts of snow, one writer has suggested, might
favor aliens with feet shaped something like skis or snowshoes. Or, creatures
in such a habitat might evolve bodies shaped like snow sleds. (It is well-known
that Antarctic penguins, with few predators around to disturb them, can travel
faster by tobogganing on their bellies than by walking.2157) An ice
planet could give rise to extraterrestrial critters with cleats or specialized
treads to improve traction.
On a flat world with few large lakes or oceans, great furry
globe-shaped creatures might be found rolling serenely across the landscape.
There are precedents for this on Earth: The tumbleweed of the American Southwest
(Sisymbrium altissimum, or tumbling mustard) and the wheel-like motions
of various spherical seeds. The extinct trilobite could curl itself into a smooth
ball capable of rolling downhill, and wood lice and certain other arthropods
retain this ability today.
Other adaptations of "undoubted utility," such
as the possibility of tractor treads in swampy environments, have been put forth
lightheartedly from time to time. The potential of rotary motion seems to cry
out for fulfillment. And yet the evolutionary process has never invented the
wheel on Earth, and Carl Sagan explains why:
Why are there no wheeled spiders or goats or elephants
rolling along the highways? Wheels are only of use when there are surfaces
to roll on. Since the Earth is a heterogeneous, bumpy place with few long,
smooth areas, there was no advantage to evolving the wheel. We can well imagine
another planet with enormous long stretches of smooth lava fields in which
wheeling organisms are abundant.15
Of course, rotary motion is not unprecedented in the animal
kingdom. We have already seen that E. coli has a tiny rotor motor to
drive its flagellae. But consider instead the significance of pearl production
in oysters, initiated by tiny grains of sand or other microscopic irritants.
Imagine an alien world the size of Earth, with continental
shelves well-flooded to a depth of tens of meters during warm spells. A creature
not unlike the molluscan cuttlefish Sepia hovers near the bottom, stalking
small fish, shrimps, and crabs. Occasionally, sand particles enter its water-jet
exit portals, clogging them up. The body responds by encasing them in perfectly
smooth spherical pearls, much like the modern oyster.
After millions of years, an Ice Age arrives and the water
slowly recede. There are few mountains due to extensive erosion, and the retreating
shoreline leaves behind vast tracts of smooth, exposed continental surface.
We might imagine our cuttlefish species, forced to live in ever shallower waters
near an increasingly turbid bottom, evolving into a kind of "caster creature."
Its several jet ports permanently plugged by large pearly
structures, such an alien might develop the ability to roll along the smooth
continental raceways. Its speed would be controlled by internal contiguous sphincters,
aided by heat sensors to allow guided braking on downhill stretches and a "low
gear" muscular assist for steep climbs. Degenerate tentacle arms could
provide additional stability on fast runs along the coastline.
The caster creature, hotly pursued by interstellar astronauts
or xenozoologists, might prove rather difficult to capture!
* Odd appendages are often used for highly
specialized tasks, as with the trunk of the elephant, the prehensile tail of
primates and the sea horse, the hind catapult bar of the spingtail insect, the
Ichneumon ovipositor and the scorpion’s massive stinger.
** Amoeboid locomotion, whether by "ectoplasmic
contraction" or protoplasmic streaming, is extremely slow. The track star
of the amoebas, Flabellula citata, can only make about 0.1 body-lengths/second
forward motion.48 Large alien amorphs probably could not better this
by much.
11.3.3
Avian Propulsion
The air is such a useful niche that it will probably be occupied
on any world with atmosphere. The ability to fly has been evolved by each of
the three major animal phyla of Earth. Among the molluscs there is a cephalopod
of the genus Omnastrephes, often called the "flying squid,"
having broad lateral flippers or fins which allow it to leap far out of the
water. Most prominent among the arthropod fliers are the insects, whose aerial
acrobats are well known.
The chordates too have many avian species. Flying fishes (e.g.,
the "flying gurnards") can leap from the water and glide hundreds
of meters before touching down. The South American Gastropelecus has
been observed to progress through the air by rapid beats of the enormous pectoral
muscles of its fins. With its keel hardly cutting the water’s surface,
this fish eventually emerges into true (but not prolonged) powered flight. The
Catalina Flying Fish is also reputed to be a strong flier.224
Reptilian gliders are widely known, including the extinct
giant pterodactyl (9-12 meter wingspan) and archaeopteryx, the flying lizards
(Draco), the so-called Flying Snake of Borneo, and so forth. Birds
are an entire class of chordates subscribing to an avian existence, and most
members of Aves can fly. There are also the gliding mammals, such as
the flying lemur and the many species of flying squirrels, and flying foxes
and bats beat the air with their wings much like birds. There are at least three
species of aerial marsupials.450
Powered flight presents a unique challenge for lifeforms on
other planets. The central problem is the need for a high power/weight ratio
-- that is, to generate enough power to get above the stall speed for the design
and into the air. (The stall speed is the slowest an object can fly and still
remain aloft.)
Typical power utilization curves for extraterrestrial avians
are shown in Figure 11.4. (The exact details of the curves are governed by something
called the Reynolds number of the wing, which relates power consumption to the
actual shape of the airfoil.) As we see, for any given wing shape and given
atmospheric conditions, there is some optimum velocity at which minimum power
is required. Flying faster or slower is less efficient and costs more energy.
Figure 11.4 Avian Power Utilization Curves2426
|
Small Avians
|
Large Avians
|
|
|
|
Power required for flight
is high during hovering (far left of each curve) and greatest at high
velocities (far right of each curve) than at the intermediate "optimum
velocity" in the trough.
Small avians such
as pigeons tend not to have serious power/weight ratio problems. Continuously
available muscular energy is sufficient to propel the animal comfortably
through a wide range of speeds (shaded area in trough). Short duration
"sprint power" permits an even wider velocity range, and is often
enough to allow jump-takeoffs from a standing start ("grasshopper
effect") or brief periods of hovering.
Large avians such
as vultures and albatrosses are not so fortunate. Their power/weight
ratio is too low to allow protracted flight even at the optimum velocity
for their wings. Hence, the large aerial animal must find a long run
way, or dive from a high perch, to reach stall speed (the minimum
for flight), and then use short duration sprint power to remain airborne
long enough to attain optimum velocity and begin relatively exertionless
soaring flight. |
If the creature flies too slowly it cannot remain aloft, and
will stall out. The speed at which this stall occurs is inversely proportional
to the square root of the density of the atmosphere.224
Given the same aerodynamic design, avians on a world with
air as thick as the atmosphere of Venus could remain airborne at speeds ten
times slower than on Earth. Conversely, on planets with Martian-thin air the
stall speed would be ten times faster than on Earth.
Stall speed is also inversely proportional to the square
root of the surface area of the wing.
An ET with huge wings can fly much slower -- and not stall
out -- than an avian with the same shape but with tiny wings. An alien with
100 m2 of wing should be able to cruise as much as ten times slower
than a creature of similar design with only 1 m2 of airfoil -- assuming
identical atmospheric conditions.
Exactly how do we go about designing an extraterrestrial
avian? How big can they be?
On Earth, the albatross is pretty close to the maximum. This
10 kg bird has been known to achieve wingspans of up to four meters. (The most
massive aerial animal that has ever lived probably weighed less than 20 kg,
although there are reports of an enormously fat cock bird shot down over the
Transvaal in 1892 which measured 24½ kg.360) The albatross
requires a lengthy "runway" for takeoff. When it lands, it must use
wing flaps like a commercial jetliner to lower the stall speed sufficiently
to land safely at about 20 kph.
The primary determinant of avian size turns out to be atmospheric
pressure, not gravity as many erroneously believe. It should also be pointed
out that the two are unrelated. High gravity does not imply high surface pressure,
as is clearly demonstrated by the members of our own solar system. For instance
Venus, our sister planet, has 9000% more pressure but 12% less gravity than
Earth.
A good empirical relation that seems to work well for most
aerodynamic lifeforms is: S = 0.24P-1(MG)0.82, where S
is total wing surface area (m2), P is air pressure (atm), M is total
body mass (kg), and G is planetary surface gravity (Earth-gees).1749
So on high pressure worlds, alien avians can make do with
vastly smaller wings. If larger wings are retained, massive bodies can be maintained
aloft. An extraterrestrial with the mass of a man, standing on the surface of
a one-gee, 5-atm world, could fly with the wings of an albatross. An albatross,
on the other hand, could make do with less than half the original wingspan.
On a 100-atm world, the 10 kg bird could be supported by stubby finlike airfoils
a mere 30 cm in breadth.
Low-pressure worlds are not amenable to large avian lifeforms
(Figure 11.5). Lift falls off rapidly, and nothing more massive than perhaps
a small pigeon would be able to take to the air. The force of gravity plays
a secondary role in fixing the size and flight characteristics of extraterrestrial
bird life. On a heavy 2.2-gee planet, the wing area would have to be about 90%
larger than an Earthly avian of comparable design. On a bantamweight 0.16-gee
world, wing area could be reduced by as much as 80% without losing the ability
to fly.
Figure 11.5 Power Requirements for
Active Flight vs. Walking Compared, for Earth-Dwellers
|
|
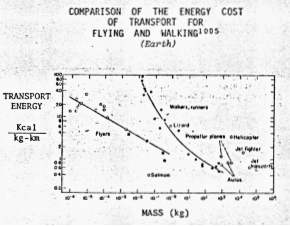
|
|
Minimum cost of transport is given both
for surface and for aerial niches. Open circles are the insect data,
squares are for birds, and filled circles are land mammals. The values
represent the coat to an animal of transporting one kilogram of its
bulk one kilometer of distance. Note that it is generally much cheaper
to fly than to walk or run, except for very large organisms.
The best-fit curves for the above data are as follows:
AIR: Cost : 1.25(MG)-0.227
LAND: Cost 10e-1,
e = 1.67(MG)-0.126
where M is total mass of the animal (kg), G is planetary surface gravity (goes), and "Cost’
is in Kcal/kg-km.
Kcal may be converted to joules by multiplying
by the factor 4190 joules/Kcal. |
|
Full line at lower right indicates the power/weight
ratio needed to sustain steady flight. Animals in the hatched area generate
insufficient power to fly.
Values are calculated on the basis of aerodynamically
optimal wing design, at the given mass on a 1-gee planet. This "minimum
power" line varies for different worlds, and in fact: P ~ 1/SQRT(r) and P ~
g where P
is the minimum power for flight,
r is the atmospheric pressure (or density),
and g is surface gravity.
Horsepower may be converted to watts by multiplying
by the factor 745.7 watts/horsepower. |
Gravity also affects the stall speed. In fact, the minimum
velocity at takeoff ~g1/2. This has several interesting consequences.
On a 2.2-gee planet the stall speed would be about 50% higher.
An extraterrestrial heavy-world albatross would require a correspondingly lengthened
runway to get off the ground.
The problem of takeoff is greatly simplified on lighter planets.
At 0.16 gees, liftoff occurs at a speed 60% below the nominal Earthly value
for the same animal. Avian ETs could perhaps take advantage of what has been
called the "grasshopper effect": An animal the size of a large pigeon
could easily hop into the minimum airspeed from a standing start on the ground.86
In light of all the evidence, we are most likely to encounter
large, intelligent winged avians on relatively small worlds with high atmospheric
surface pressures.
How many wings are best?
The first winged insects on ancient Earth probably had no
more than three pairs. A few modern insects retain vestigial traces of the third
pair, notably the Stenodictya of the Order Strepsiptera.1212
By and large, however, aerial arthropods have cut down on the number of wings,
Locusts, ant lions, fishflies, termites and aphids each have two pairs apiece,
but a single pair is far more common in the animal world,
There are good reasons to reduce the wing count. The usual
arrangement is a single pair, which serves more or less in the capacity of a
helicopter rotor -- that is, to generate active lift. Another less common design
is to use one pair of wings to generate passive lift (like the wings of an airplane)
and a second pair taking the more active role. (Beetles come close to doing
just this.) But no useful purpose is served by adding more wings, which would
only interfere with the smooth airflow and ruin the aerodynamics of the design.1212
Using the principle of economy, extraterrestrial avians will
have one pair, or at most two pairs, of wings.
Or they may have no wings at all. Thus far we have discussed
only the most common techniques of flight known on Earth. But there are many
other -- wingless -- ways to get into the air. On this planet, the principles
of the rocket, the kite, and the balloon have not been widely exploited.
Imagine a world with a thick atmosphere rich in oxygen and
abundant seas. Evolution might favor large but slow-moving insects as discussed
earlier in connection with respiration. We might expect a kind of "rocket
fish" to arise near the coasts, feeding where such insect life swarmed
thickly in the air.
Like the small plastic toy projectiles that shoot high into
the air when fully charged with water and compressed gas, the rocket fish suddenly
would bolt from the sea skyward and mouth their aerial prey at high speeds.
To be able to eat on the run, this jet-propelled alien predator has to evolve
a sturdy posterior pressure canister which can be discharged rapidly through
a rigid nozzle. The rocket fish might have a recycling time on the order of
minutes, and might charge their canisters using osmotic pressure.
Another class of alien creatures might take to the air on
other worlds. A low gravity planet with fast rotation and a thick atmosphere
could be ideal for the evolution of "parachute beasts." These ETs
could travel virtually anywhere for free, simply by extending their retractable
‘chutes. With strong winds, they could tack across the planetary surface.
With a good stiff breeze, even fairly massive parachute beasts could kite hundreds
of kilometers each day without effort.
There is some precedent among Earthly fauna. In the majority
of spider species, aerial dispersal of the young takes place. A spiderling crawls
to the end of a blade of grass or other protuberance, raises its tiny abdomen,
and lets fly a thin thread of silk into the wind. As the gossamer strand is
caught by a breeze, the spiderling leaps from its perch and climbs to the middle
of this floating "magic carpet." Air currents carry the animal to
considerable distance: Spider threads have been sighted at least as high as
8 km and as far seaward as 500 km from shore, Couldn’t extraterrestrial
"hang gliders" do just as well?
The idea of the balloon principle in relation to living organisms
is a common one, both in science and science fiction.20,442,2427
It appears, for instance, in a nineteenth-century novel by the French writer
Charles Ischir Defontenay -- Psi Cassiopeia (1854):
Here is what the Starian naturalists say of this animal,
which they call the psargino: Its skin, which has great extensibility, is
only attached at the eyes, the mouth, the other natural openings of the body
and the soles of the feet. Over the rest of its expanse, it is only juxtaposed
to another membrane or internal skin having the property of secreting, at
the animal’s will, {hydrogen} gas. The psargino, thus surrounded by
gas, becomes a sort of balloon lighter than the atmosphere, and it makes use
of this property to rise into the air and escape its enemies. A kind of aperture
furnished with valvules on its abdomen relieves it of part or all of the gas
burdening it and serves for descent to earth when the predator has lost its
track.564
Bonnie Dalzell has designed an "airship beast"
for a planet with cold winters, heavy gravity and a thick atmosphere.736
Twice a year this herbivorous 100 kg animal inflates its lifting bags with metabolically
generated hydrogen gas* and drifts to the opposite hemisphere to avoid the cold.
Strong winds are an advantage, but predators are widespread. And many of these
living balloons are lost during their semiannual migration when lightning from
an electrical storm strikes and ignites their bodies.
There are a few indirect precedents for such a creature among
the lifeforms of our own world.2574 For example, the Portuguese man-of-war
(Physalia) is a shapeless, baglike marine organism whose large air
sac serves both as a float and a sail in the water, Its numerous tentacles perform
many functions, including stinging its prey into senselessness,
Another creature even more analogous to the airship beast
is the chambered nautilus. Some 3000 species of this animal once flourished
in the primitive, shallow seas of our planet. They are found as deep as 700
meters, and although they measure a mere 0.2 meters today their ancestral forebears
left fossil remains up to three meters in diameter.584
The nautilus is a miniature submarine, consisting of a series
of as many as forty individual chambers partially filled with air. All are connected
by a thin tube called the siphuncle, which is thought to control the buoyancy
of the animal as it dives and ascends in water.
As the man-of-war and the chambered nautilus utilize the
principle of buoyant lift in the seas of Earth, why could not giant living extraterrestrial
gasbags ply the skies of alien worlds?
* Many bacterial photosynthetic autotrophs
generate hydrogen metabolically. Examples include Clostridium, Chromatium,
Athiorhodaceae, and green algae under certain special circumstances.
Back to Contents