|
from Hindawi Website
The radio frequency (RF) environment of the earth has undergone major changes in the past two centuries due to the development and use of electricity in power and communications applications, and the anthropogenic RF background continues to increase in intensity and complexity.
This study suggests that the RF
background may have strong adverse effects on growth rate and fall
anthocyanin production in aspen, and may be an underlying factor in
aspen decline.
Stand age and clonal differences are also factors in susceptibility to decline [6]. Factors initiating decline include: defoliation by insects, damage caused by wildlife, severe drought, and extreme weather incidents [7]. Damage caused by these factors can diminish the vigor of affected clones and make them vulnerable to opportunistic fungal pathogens and insects [8].
The concept of forest decline has been used to describe the interaction of these various factors; however, the underlying causes of aspen decline are not well understood [9]. Since 2004, incidents of very rapid aspen clone mortality have been seen in Colorado [10].
Because the electromagnetic environment (EM) of the earth has changed radically in the past two
centuries, this study investigates the possibility that
anthropogenic changes in this environment, particularly in the radio
frequency (RF) spectrum, are adversely affecting growth and health
of aspen populations, making them vulnerable to decline.
The source of the EM input, however, makes no difference. Timing of plant processes is an important mechanism for plant protection and efficient functioning in changing day/night and seasonal environmental conditions [11].
Although photosynthesis requires fairly strong energy input in the blue and red visual frequencies (full sun/shade) [12], photoperiodic responses in plants are typically triggered by energy inputs in the red and far-red frequencies that are in the range of 10−4 times the energy required for photosynthesis, and even a brief flash of light during a plant’s subjective night can be enough to trigger a short night response, strongly affecting plant behavior and morphology [13].
The radio frequencies, lower than 300 gigahertz, are below visual and infrared frequencies in the electromagnetic spectrum. The earth’s natural RF environment has a complex periodicity that has been more or less the same within the lifespan of modern tree taxa. Before 1800, the major components of this environment were broadband radio noise from space (galactic noise), from lightning (atmospheric noise), and a smaller RF component from the sun [14].
Because of the periodic nature of the naturally occurring RF background, plants may have evolved to use those environmental signals, as well as visible light, to regulate periodic functions, and therefore they may be sensitive to anthropogenic RF input. The intensity of the human-generated RF environment has increased gradually since about 1800.
This background of RF pollution is now
many times stronger than the naturally occurring RF environment.
From the perspective of evolutionary time, the change can be
considered sudden and dramatic [14, 15].
However, the assumption seems to have been made that the low-level background of RF pollution has no effect on biological systems. Therefore, not only has this particular phenomenon not been studied, but it has been generally disregarded in earlier experiments designed to look for effects of RF exposure. Results of those previous experiments are often difficult to duplicate, and it may be that the anthropogenic RF background has been a confounding factor.
However, for some RF effects on biological systems, consistent results have been documented in previous experiments:
This experiment investigates the effects of the radio frequency background on aspen seedlings.
The objective of this preliminary study
was to determine if aspens respond to the current RF background in
ways that result in reduced growth, or in increased susceptibility
to pathogens.
1.1. Study Area
The ecological system in the area of the experiment is Rocky Mountain Lower Montane. The site was about 400 meters below the lower elevation of aspen’s natural range in Colorado. The north-facing slope was chosen as a somewhat cooler microclimate than other aspects. Vegetation included various grasses, which were mowed on the 8×10m experimental plot.
Vegetation in the east, south, and west directions from the experiment included:
North of the experiment, there were
large cottonwood trees along the river, with grass and chokecherry
undergrowth.
1.2. Materials and Methods
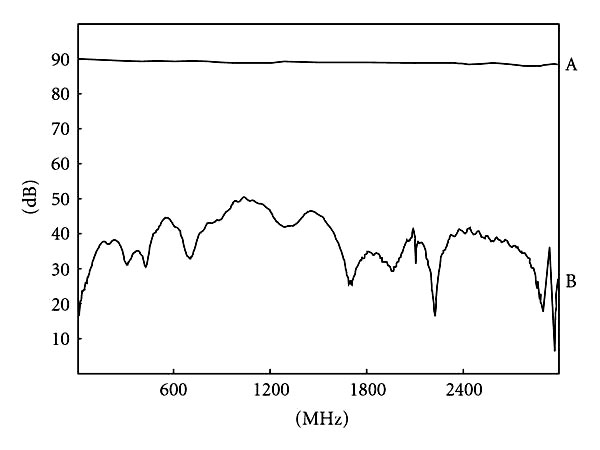
The aluminum screen material in the configuration used for this experiment was tested at a laboratory of the National Institute of Standards and Technology (NIST) in Boulder, CO. From 1.0 MHz to 3.0 GHz, its effectiveness in attenuating RF signals was found to vary from 40 dB to 73 dB across the entire range of frequencies (Figure 1).
Theoretically, an enclosure made of this material would reduce signal intensity by a factor of 10-4 to 10-7.3, blocking essentially all ambient RF energy, including the naturally occurring RF background. In practice, the shielding effectiveness of a Faraday cage made of this material could be somewhat less than that of the material itself due to gaps or imperfect electrical contact at seams and closures, and to the internal RF acoustical properties of the cage.
A similar cage was constructed as a control, with fiberglass screen, which is not conductive and which does not block RF signals, instead of aluminum. A transistor radio was used to test the effectiveness of the cages to block the RF background. No radio stations could be picked up inside the Faraday cage.
In both the mock-Faraday cage and the uncaged area, numerous stations were received in the AM and FM bands.
A sweep of RF background at the site, June 6, 2009, using an Anritsu spectrum analyzer, showed that field intensity ranged from -117 dBm to -87 dBm at frequencies from 1 to 1,000 MHz. Mean field intensity was -109 dBm.
The aluminum screen material in the configuration used for this experiment was tested at NIST in Boulder, CO. From 1.0 MHz to 3.0 GHz, its effectiveness in blocking RF signals was found to vary from 40 dB to 73 dB across the entire range of frequencies. X axis = Radio frequency from 1.000 MHz to 3000.000 MHZ in increments of 300.000 MHz left to right. Y axis = Decibel scale. Reference signal level was 90 dB. A = Reference signal. B = Attenuated signal.
Seed for these seedlings came from Caffey County near Salida Colorado, approximately 195 km SSW of the experiment site, at an elevation of about 2,100 meters. The seeds were germinated in June 2006 at the CSFS Nursery, and the resulting seedlings were grown in a shade house in a Styrofoam planter-block of 30 seedling plugs.
The seedlings had leafed out in early spring and, when purchased on May 28, 2007, were not producing new leaves. Seedlings were transplanted into no. 1 black gallon plastic pots on June 5, 2007 using a commercial potting medium, Black Gold, composed of: 45–55% Canadian sphagnum peat moss, compost, pumice, perlite, and composted worm castings (N 0.05%: water soluble 0.006%, water insoluble 0.044%).
Since only 27 seedlings were needed for the experiment, due to space restrictions, the three smallest seedlings were discarded.
The remaining seedlings were relatively uniform in height, stem diameter, and leaf development, and the leaves appeared healthy and green. The 27 seedlings were assigned to 3 groups of 9 seedlings each, with variation in size distribution distributed equally among the 3 groups.
After potting, the three groups were photographed and randomly assigned to the mock-shielded cage, the shielded cage, or to the unshielded area. The experiment began June 6, 2007. All treatments in the experiment were exposed to full sun from sunrise to late afternoon. There was a distance of three meters between treatment enclosures. All seedlings in the experiment were watered at the same time 1-2× per week, depending on the weather, using Lyons water.
Seedlings were watered with a weak
solution of fertilizer (5-10-5) on July 29, 2007, and otherwise were
not fertilized.
The monitoring devices were placed in the middle of the monitored treatment enclosure, just above the top of the pots. A board was placed south of the devices to provide shade. A lux meter was used to measure light intensity in the treatment areas. Shielding reduced light intensity by 35% for the mock-shielded enclosure and 40% for the RF-shielded enclosure.
Naturally occurring sunlight intensity was not reduced for the group that was not shielded. Except for the difference in RF background intensity, conditions in the shielded and mock-shielded enclosures were very similar.
The unshielded seedlings were exposed to higher light levels (full sun), higher airflow, and generally lower humidity than the shielded and mock-shielded treatments since they were not in a screened enclosure.
Because of that, there was a shoot node dividing spring growth from the summer growth phase that was easy to identify. Measurement of active leader (shoot) length was taken from the tip of the shoot down to that first shoot node, so leader length measured was only that produced after the experiment began on June 6, 2007.
For leaves emerging directly from a main
stem, leader length was recorded as 0.00 cm.
After measurements were taken, all seedlings were returned to their cages.
October 5-6 photos were taken showing differences in leaf coloration, and relative leaf area affected by necrotic lesions.
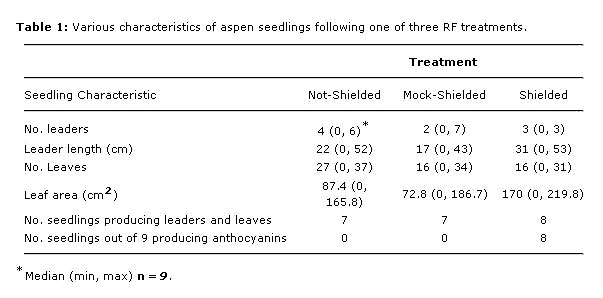
The RF shielded group evidenced more vigorous growth, producing 74% more total leader length and 60% more leaf area than the mock-shielded group, though the number of leaves produced by both groups was nearly the same (Table 1).
Various characteristics of aspen seedlings following one of three RF treatments.
The two RF-exposed groups’ morphology and behavior were very similar and within the current norm for aspen, including absence of fall anthocyanin production, and extensive leaf area affected by necrotic lesions in fall senescing leaves.
On active leaders, the shielded group showed fall production of anthocyanins far outside the norm, absence of fall leaf tissue necrotic lesions, and a wide range of fall colors:
Shielded leaves produced in the spring flush (before treatment), however, were similar in appearance to leaves in the two exposed groups in color, size, and incidence of lesions.
All of the shielded leaves on active leaders had dark to bright red veins and petioles indicating strong anthocyanin production (Figure 2).
On October 5, 2007, unshielded and mock-shielded seedlings had leaf tissue ranging in color from yellow to green, and a high percentage of leaf tissue in both exposed groups displayed necrotic lesions (Figures 3 and 4).
Shielded seedlings had larger leaves, petioles and leaf veins were red, and tissue colors varied from yellow to dark green (Figure 5). Some shielded leaves looked black because of the combination of red veins and dark green tissue (Figure 6).
Leaves on active leaders in the shielded group were substantially free of leaf tissue lesions (Figure 7), but unshielded and mock-shielded leaves were all affected to some extent by leaf tissue necrosis (Figure 8).
October 6, 2007. Mock-shielded seedlings on the left and shielded seedlings on the right,
the latter showing
more total leaf area, stronger fall leaf coloration, and minimal
leaf necrosis symptoms.
Unshielded seedling, showing green to yellow leaf coloration
and gray to brown
necrotic leaf tissue, Oct. 6, 2007.
Mock-shielded seedling,
exhibiting yellow
leaf coloration and brown lesions, Oct. 6, 2007.
Shielded seedling, showing red petioles and veins on light green leaf tissue,
and very little leaf
tissue affected by necrosis. Oct. 6, 2007.
Shielded seedling. Combination of red veins and green leaf tissue
makes leaves look
black (left side of photo), Oct. 6, 2007.
Shielded seedling, with leaves on active leaders mainly free of necrotic lesions,
and leaf veins and
petioles red, Oct. 6, 2007.
Mock-shielded seedling, showing all leaves affected by necrotic lesions to some extent, leaf veins yellow or green, and petioles light red to pink, Oct. 6, 2007.
3. Discussion
Results of this experiment do, however, suggest several possible effects of RF exposure on aspen. Those effects include reduction of shoot length and leaf area, suppression of fall anthocyanin production, and increased incidence of necrotic leaf tissue in fall senescing leaves.
Reduced vigor in the two RF-exposed groups is demonstrated by lower total leader length and diminished production of leaf area relative to the shielded group.
A study [26] comparing mutant nonanthocyanin-producing strains of fall anthocyanin-producing species, Cornus sericea, Vaccinium elliottii, and Viburnum sargentii, showed very similar results to this experiment with regard to fall color in that extensive necrotic leaf tissue was seen in nonanthocyanin-producing mutants during fall leaf senescence and leaves dropped while still green.
Wild-type strains of those species, which produced anthocyanins, did not exhibit necrotic lesions. In that study, leaf tissue damage in mutant strains was shown to be caused by photooxidative stress. During fall leaf senescence photosynthetic mechanisms become vulnerable to damage by visible light. Anthocyanins shield the photosynthetic apparatus from high light levels, preventing photoinhibition and damage to leaf tissue due to light stress.
Transport of foliar nutrients requires
energy provided by photosynthesis [27], and since the
majority of nutrients in overwintering deciduous trees are derived
from foliar nutrient transport during autumn leaf senescence,
differences in anthocyanin production could have major effects on
plant fitness [28].
Although shielding materials
substantially reduced light levels in the two caged groups
(mock-shielded and shielded), the behavior and appearance of the two
RF-exposed groups (mock-shielded and unshielded) were most similar,
both showing no apparent anthocyanin production, leaf tissue
remaining light green or yellow, and a high percentage of leaf
tissue affected by necrotic lesions.
3.1. Radio Frequency Environment
Currently, the RF spectrum is quite crowded, and new technologies are constantly being developed to allow more information to be transmitted on the available frequencies [29].
Unintended broadband RF noise is created as a by-product of electrical power generation, transmission, and use. These human-made RF sources create a background noise (as differentiated from signal) level generally 10 to 100 times stronger than the naturally occurring background of galactic noise and atmospheric noise at frequencies below 30 MHz [14].
Human-generated signal intensities in large regions of the RF spectrum are in the range of 103–106 times stronger than the naturally occurring RF background in urban areas [15].
Because terrestrial and satellite RF sources are numerous and widely dispersed globally, and since below 30 MHz radio signals can travel great distances around the earth by bouncing between the ionosphere and the earth’s surface [14], currently a strong human-generated RF background exists at every point on the earth’s surface, although radio field strength is relatively greater in the most populous and urbanized areas.
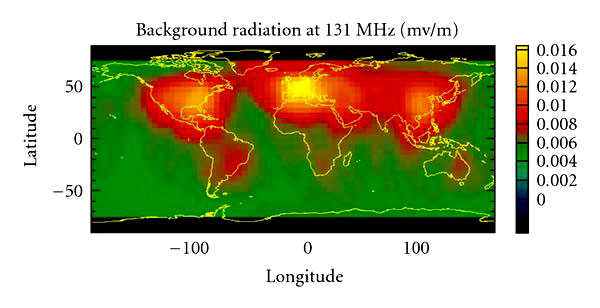
Globally, the highest field strengths occur in central Europe, the eastern United States, and in China (Figure 9).
Forest decline was first recognized and defined based on observed events in central Europe and the eastern US, and China, at this time, is experiencing rapid desertification.
Forte Satellite: 131 MHz RF background 2000. Areas of highest radio background intensity occur in the eastern United States, central Europe, and China.
Acquired by the Los
Alamos National Laboratory FORTE spacecraft. Principal investigator
A.R. Jacobson.
3.2. Forest Decline
In a European study [31], stress factors related to forest defoliation were found to be age, altitude, drought, and various types of air pollution, in descending order of importance.
Although the statistical correlation was high for some of these effects, because the number of samples was large, the effects documented were quite small and altogether could account for only 15-55% of the observed decline, depending on species. No temperature variables were found to have significant effects. In several studies, climate change was found not to be a causal agent in forest decline [32, 33].
There is evidence, however, that trees involved in forest decline are less tolerant of extreme weather conditions.
Freeze damage, possibly indicating disrupted dormancy, for instance, has been noted as one symptom of forest decline where temperature and/or precipitation pattern changes were not evident [32, 34].
More recently, it has been shown that mortality rates of all dominant tree species in the western United States have been doubling every 17-29 years in old growth forests, and that recruitment of new trees is now occurring at a lower rate than mortality [35].
Since aspen decline and other tree
decline incidents worldwide have similar symptoms, and since no
definitive explanation has been found for those events, it seems
plausible that their decline may be related to RF exposure.
Changes would have occurred very slowly over time, from the human perspective, and would have been widely dispersed geographically so that, at any particular time, RF-altered plant morphology and behavior would have been normal, the norm, by definition.
Plants and animals have in previous experiments been shown to respond in various ways to RF exposure.
Growth of organisms may be inhibited or accelerated by RF exposure, thus possibly affecting interactions at the level of communities and ecosystems from microscopic to global scales.
In the case of aspen, reduced plant
vigor and adverse effects on fall storage of nutrients due to RF
exposure could leave seedlings vulnerable to pathogens and less able
to accommodate other environmental stress.
4. Conclusions
These effects suggest that exposure to the RF background may be an underlying factor in the recent rapid decline of aspen populations. Further studies are underway to test this hypothesis in a more rigorous way.
References
|