|
by Christine M. Kaefer and John A. Milner Chapter 17 of "Herbal Medicine - Biomolecular and Clinical Aspects. 2nd edition" 2011 from NCBI Website
17.1 - INTRODUCTION
Historically, spices have shaped many events throughout the world.
Many voyagers, including the legendary Christopher Columbus, explored the seas in search of treasured spices. These valued commodities contribute not only flavors but also serve as colorants and preservatives in a wide variety of cultures.
Today, spices are increasingly revered not only for their culinary properties but also for their potential health benefits.
Although the health attributes associated with spice use may arise from their antioxidant properties, their biological effects may arise from their ability to induce changes in a number of cellular processes, including those involved with drug metabolism, cell division, apoptosis, differentiation, and immunocompetence.
The complexity of understanding the biological response to spices first surfaces in the criteria used to distinguish what constitutes a culinary spice and how they differ from culinary herbs. These terms are often used interchangeably in the scientific and lay literature.
The U.S. Food and Drug Administration (FDA) defines a spice as an,
While this is a viable definition, it does not consider the biological consequences of consuming these items and how they differ from herbs.
The U.S. National Arboretum offers an alternative definition and describes spices as,
We must remember that the quantity of an item consumed does not dictate its importance.
Thus, to avoid the health significance in any definition would appear flawed. In this chapter, we use the terms "herbs" and "spices" interchangeably and assume that both have properties that extend beyond simply providing flavor and color.
There is little doubt that nutrition and health are intimately linked (Kennedy 2008).
For generations, people have alleged that foods provide greater benefits than simply supplying energy. Beliefs in the medicinal properties of foods have surfaced in many early writings of man.
Hippocrates is frequently quoted as having said "Let food be thy medicine and medicine be thy food." Epidemiological, preclinical, and clinical studies continue to provide fundamental insights into the dynamic relationships between nutrients - defined here as any substance in the diet that brings about a physiological effect - and health.
Today, claims about the ability of foods, including spices, to lower disease risk or to enhance the quality of life continue to captivate our lives (Kaefer and Milner 2008; Kochhar 2008; Krishnaswamy 2008; Iyer et al. 2009).
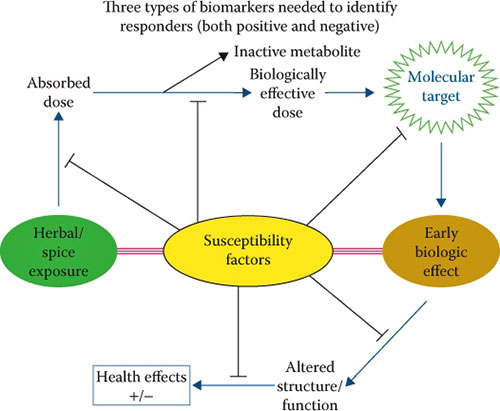
Three types of biomarkers - exposure, effect, and susceptibility - are needed to evaluate the effects of spices in cancer prevention and therapy (Figure 17.1).
Additional information about the amounts of specific spices required to bring about a response (effect) and the interactions of spices with other constituents of the diet, microbes in the gastrointestinal tract, environmental exposures, and human genetics (susceptibility factors) will be needed to unravel the true benefits of adding spices to the diet.
FIGURE 17.1(See color insert.) Three types of biomarkers (exposure, effect, and susceptibility) are needed to assess the benefits or risk of spices.
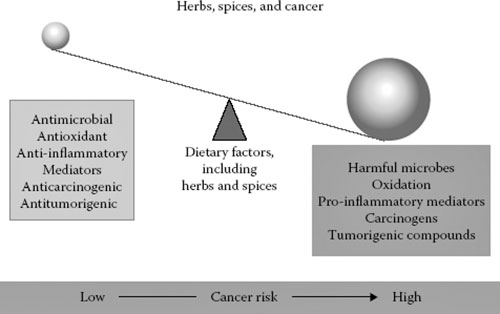
Spices may be a key to determining the balance between pro- and anticancer factors that regulate risk and tumor behavior (Figure 17.2).
About 75% of U.S. households use dietary approaches to reduce their risk of diseases, including cancer (Sloan 2005).
Americans between the ages of 36 and 55 are increasingly interested in adopting healthy eating behaviors and are gravitating toward ethnic cuisines based on perceived health benefits (Uhl 2000).
Many of these ethnic foods are loaded with unique and flavorful spices; however, while dietary guidelines in several countries tend to support the incorporation of spices into diets, quantifiable recommendations for specific amounts have not yet been forthcoming (Tapsell et al. 2006).
FIGURE 17.2Multiple factors may influence the need for spices for reducing the risk of cancer or changing the biological behavior of cancerous cells.
Between 1970 and 2005, the overall per capita consumption of spices in the United States doubled, increasing from about 1.6 to 3.3 pounds per year (United States Department of Agriculture Economic Research Service, 2007).
As expected, the consumption of some spices increased far more than others; for example, garlic consumption increased more than sixfold.
According to a report by Buzzanell (1995) from the United States Department of Agriculture’s (USDA) Economic Research Service, the increasing domestic use of spices reflects a number of factors. Among these are the growing Hispanic and Asian populations within the United States, an increasing trend toward the use of culinary herbs and spices to compensate for less salt and lower fat foods and a general increase in the popularity of ethnic foods.
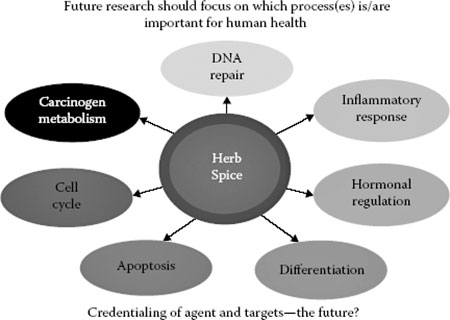
This chapter reviews culinary herbs and spices for their ability to modify several cellular processes that are linked to the risk of cancer and/or tumor behavior (Figure 17.3).
The ability of spices to serve as inhibitors of carcinogen bioactivation, decrease free radical formation, suppress cell division and promote apoptosis in cancerous cells, suppress microbial growth, and regulate inflammation and immunocompetence will be discussed as plausible mechanisms by which selected spices may promote health and disease resistance.
The low toxicity and wide acceptance of spices may make them particularly useful as a subtle personal dietary change that may decrease risk for several diseases. It is already appreciated that the addition of about 1 g/day of herbs to one’s diet can significantly contribute to total antioxidant intake (>1 mmol) and offers a better source of antioxidants than many food items (Dragland et al. 2003; see also Chapter 2 on antioxidants in herbs and spices).
Because several spices are effective antioxidants, they may be particularly important in decreasing oxidative damage due to environmental stress, including excess calorie intake.
FIGURE 17.3Multiple cancer-related processes may account for the ability of spices to inhibit experimentally induced cancers. While these processes are likely critical for determining the risk of cancer and tumor behavior in humans, only limited clinical evidence exists that spices in physiological relevant exposures can alter one or more of these process.
More than 180 spice-derived compounds have been identified and explored for their health benefits (Aggarwal et al. 2008).
It is beyond the scope of this chapter to deal with all herbs and spices that may influence the risk of cancer and tumor behavior.
Therefore, a decision was made to review those with some of the more impressive biological responses reported in the literature, and a conscious effort was made to provide information about the amount of spices needed to bring about a response and thus their physiological relevance.
When possible, recent reviews are included to provide readers with additional insights into the biological response(s) to specific spices and to prevent duplication of the scientific literature.
Because there is a separate chapter devoted to curcumin (a bioactive component in turmeric) in this book and there are also several excellent reviews published about curcumin (Patel and Majumdar 2009; Aggarwal 2010; Bar-Sela, Epelbaum, and Schaffer 2010; Epstein, Sanderson, and Macdonald 2010), turmeric is not discussed in this chapter.
17.2 - ALLSPICE
The term "allspice" was coined in the 1600s by the English, who thought the herb combined the flavors of cinnamon, nutmeg, and cloves.
Allspice is also referred to as,
Ground allspice is not a mixture of spices as some still believe, but arises from the dried unripe berries of the tree Pimenta dioica.
This tree is native to the Greater Antilles, southern Mexico, and Central America. Today, P. dioica is cultivated in many warm areas throughout the world. Allspice is also available commercially as an essential oil.
Allspice is claimed to possess properties such as,
It contains a multitude of potential bioactive agents that may contribute to health promotion, including flavonoids, phenolic acids, catechins, and several phenylpropanoids (Al-Rehaily et al. 2002).
Berries contain about 2-5% essential oils that include the following bioactive compounds: eugenol (60-75%), eugenol methyl ether, cineole (eucalyptol), phellandrene, and caryophyllenes (Kluth et al. 2007).
The antioxidant and antimicrobial activities of allspice may be associated with eugenol (Rompelberg et al. 1996; Kluth et al. 2007).
Billing and Sherman (1998) reported that allspice was as effective as garlic and onions in suppressing microbial growth. The significance of its antimicrobial properties was recently highlighted by evidence that allspice and eugenol were effective in lowering the virulence of Escherichia coli O157:H7 (Takemasa et al. 2009).
Nevertheless, there are concerns that allspice oil can be toxic and promote inflammation, nausea, and vomiting when consumed in excess.
The anticancer properties of allspice may be in part due to its ability to influence cytochrome P450 (CYP) activity and thereby influence carcinogen bioactivation. Kluth et al. (2007) cultured human liver carcinoma cells and human colon adenocarcinoma cells and studied the ability of the spice extract to activate mechanisms related to phase I detoxification enzymes.
The allspice extract (3 mg/mL in dimethyl sulfoxide) did not activate pregnane X receptor (PXR) directly but did strongly activate the CYP3A4 promoter. Thus, the activation of transcription factors to bind to response elements seems like a plausible mechanism by which allspice, and potentially eugenol, function.
There is specificity in the response to allspice and eugenol because gastrointestinal glutathione peroxidase (GPx), a phase II enzyme linked to removal of reactive oxygen species (ROS), was not influenced by allspice or eugenol (Kluth et al. 2007).
Inflammation is linked to increased risk of cancer (Dinarello 2010) and appears to be influenced by allspice consumption. Although controlled clinical interventions are not available, evidence in rodents suggests potency (Al-Rehaily et al. 2002).
Providing an oral allspice suspension (500 mg/kg body weight) significantly inhibited carrageenan-induced paw edema and cotton pellet granuloma in rats.
It also suppressed acetic acid-induced writhing and tail flick reaction time and decreased yeast-induced hyperpyrexia in mice. Interestingly, the suspension also appeared to have antiulcer and cytoprotective activity in rats by protecting gastric mucosa against indomethacin and various necrotizing agents, including 80% ethanol, 0.2 M sodium hydroxide (NaOH), and 25% sodium chloride (NaCl), suggesting that it might also have an impact on cyclooxygenase (COX) activity.
It remains unclear what molecular target alteration(s) account for this response.
Evidence exists that allspice can alter the proliferation of several cultured cancerous cells. While cell viability was reduced about 50% when allspice extract was added to prostate cancer cells (LNCaP cells), it did not influence the viability of cultured human prostate cancer cell lines (DU145) or cervical epithelial carcinoma (HeLa) cells (Lee et al. 2007).
The mechanism by which allspice leads to cellular growth depression remains largely unresolved. However, recent studies by Lee et al. (2007) suggest that epigenetics may be involved.
A depression in histone acetyltransferase (HAT) activity may be involved. Androgen-induced HAT activity was decreased by 70% when allspice was provided at 100 μg/mL. Allspice also suppressed androgen receptor (AR) acetylation in LNCaP cells and significantly decreased histone H3 and H4 acetylation, indicating that a repression of AR-mediated transcription was induced due to shifts in histone and nonhistone acetylation.
While these in vitro studies are intriguing, there is a need for controlled interventions in animal models before exploring allspice’s potential benefit as a dietary antitumorigenic agent.
17.3 - BASIL
Basil (Ocimum basilicum) is a culinary herb prominently featured in Italian and Southeast Asian cuisines.
While many varieties of basil exist, sweet basil is one of the most predominant and most frequently examined herbs for its health benefits. Basil is originally native to Iran, India, and other tropical regions of Asia, but now it is widely available throughout the world.
Basil’s antioxidant, antimutagenic, antitumorigenic, antiviral, and antibacterial properties likely arise from a variety of components including linalool, 1,8-cineole, estragole, and eugenol (Muller et al. 1994; Chiang et al. 2005; Makri and Kintzios 2007).
Similar to most culinary spices, far more information is needed about the variation in content of constituents as a function of plant varietal, growing conditions, and processing.
The essential oil of basil possesses antimicrobial properties (Wannissorn et al. 2005). Moghaddam, Karamoddin, and Ramezani (2009) investigated the effect of basil on Helicobacter pylori and found that methanol, butanol, and n-hexane fractions of basil demonstrated antagonistic activity against the bacteria (MIC = 39-117 μg/disk).
While not as potent as amoxicillin, its effectiveness raises possibilities of using individual or multiple spices as potent antimicrobials, especially in areas where commercial antibiotics are in limited supply (Moghaddam, Karamoddin, and Ramezani 2009).
The effects of basil are not limited to its antimicrobial properties because evidence indicates that it also can lower oxidative damage in animal models (Dasgupta, Rao, and Yadava 2004).
Feeding mice 200 and 400 mg/kg body weight with a hydroalcoholic extract of basil leaves for 15 days markedly increased GPx (1.22-1.4 fold), glutathione (GSH) reductase (1.16-1.28 fold), catalase (1.56-1.58 fold), and superoxide dismutase (1.1-1.4 fold; Dasgupta, Rao, and Yadava 2004).
The change in activity in one or more of these enzymes may explain the decrease in lipid peroxidation caused by basil in studies by Dasgupta, Rao, and Yadava (2004). Drăgan et al. (2007) examined the effects of balsamic vinegar–enriched extracts from several herbs (rosemary, sage, and basil) in soups and salads on oxidative stress and quality of life measures in women with stage IIIB and IV breast cancer.
While there was a decrease in oxidative stress, the complexity of the dietary intervention made it impossible to determine the component(s) that led to improvements.
Several studies provide evidence that basil is an antimutagenic spice (Kusamran, Tepsuwan, and Kupradinun 1998; Stajkovic et al. 2007). Stajkovic et al. (2007) studied the antimutagenic effects of basil on mutagenicity in Salmonella typhimurium TA98, TA100, and TA102 cells in the presence or absence of liver microsomal activation.
The essential oil of basil, at concentrations ranging from 0.5 μL/plate to 2.0 μL/plate, inhibited mutations from ultraviolet irradiation (dose = 6 J/m2) by 22-76%.
Mutations caused by 4-nitroquinoline-N-oxide (0.15 μg/plate) were decreased by 23-52%, and those from 2-nitropropane (14.9 mg/plate) by 8-30%. These findings are consistent with studies by Jeurissen et al. (2008), who demonstrated that 50 μg/mL basil largely blocked DNA adduct formation caused by 1′-hydroxyestragole in the human hepatoma (HepG2) cell line, possibly by promoting phase II enzymes and thereby conjugation and elimination of this carcinogen.
These findings likely explain the ability of basil to decrease the mutagenicity of aflatoxin B1 (AFB1) and benzo(a)pyrene (B(a)P) (Stajkovic et al. 2007).
The mutagenicity of AFB1 was inhibited by >30% by the presence of 1-2 mg/plate of a hexane-based basil extract and 0.5-1 mg/ plate of chloroform- and methanol-based basil extracts. Because B(a)P mutagenicity was only inhibited by chloroform- and methanol-based basil extracts at doses of 2-5 mg/plate, multiple constituents might be responsible for basil’s antimutagenic activities.
The anticancer properties of basil in preclinical studies are mixed. In studies with Sprague-Dawley rats fed with an AIN-76 diet with or without high concentrations of basil (6.25% and 12.5%), there was no clear indication of a decrease in 9,10-dimethyl-1,2-benzathracene (DMBA)-induced mammary cancer.
It is unclear whether the quantity of the procarcinogen examined, the simultaneous induction of both phase I and II enzymes, or some other factors accounted for the lack of protection by adding basil to the animals’ diet (Kusamran, Tepsuwan, and Kupradinun 1998). Nevertheless, there is evidence that basil can decrease DMBA-induced carcinogenesis.
Providing Swiss mice with a diet containing 150 or 300 mg/kg body weight of basil extract decreased DMBA-induced skin tumors (12.5% reduction and 18.75% reduction for lower and higher doses, respectively), and lowered the tumor burden per mouse.
Compared to the average number of tumors per mouse in the controls, the tumor burden was approximately 2.4 times lower (p < .01) in the low-dose basil group and 4.6 times lower (p < .001) in the high-dose basil group (Dasgupta, Rao, and Yadava 2004). It is unclear whether differences in the response between mice and rats reflect the species, the cancer site, or the dietary or procarcinogen exposures.
DNA methyltransferase (MGMT) is a critical repair protein in the cellular defense against alkylation damage. MGMT is highly expressed in human cancers and in tumors resistant to many anticancer alkylating agents. Niture, Rao, and Srivenugopal (2006) examined the ability of several medicinal plants to upregulate O6-methylguanine adducts.
Both ethanol and aqueous extracts of basil increased MGMT protein levels in HT29 human colon carcinoma cell lines 1.25-fold compared to controls after 72-hours incubation. Compared to the control, basil increased glutathione-S-transferase (GST) protein activity 1.33-fold after 12 hours of incubation; after 24 hours, GST activity increased 1.68-fold compared to the control, which declined to 1.47-fold after 72 hours incubation.
Because MGMT is one of the body’s first lines of defense against alkylation DNA damage, a small increase (two- to threefold) in this enzyme may protect against mutagenic lesions (Niture, Rao, and Srivenugopal 2006).
The anticancer properties of basil may also relate to its ability to influence viral infections. Individuals with hepatitis B are recognized to be at increased risk for hepatocellular carcinoma (Fung, Lai, and Yuen 2009; Ishikawa 2010). Chiang et al. (2005) evaluated the antiviral activities of basil extract and selected basil constituents in a human skin basal cell carcinoma cell line (BCC-1/ KMC) and a cell line derived from hepatoblastoma HepG2 cells (2.2.15) against several viruses, including hepatitis B.
Impressively, Chiang et al. (2005) found that the aqueous extract of basil, along with apigenin and ursolic acid, displayed greater anti-hepatitis B activity than two commercially available drugs, glycyrrhizin and lamivudine (3TC).
Overall, these studies raise intriguing questions about the merits of using commercially available spices to retard viruses and potentially cancer. Undeniably, much more information is needed to clarify the amounts and durations needed to bring about a desired viral response and the mechanism by which a response occurs.
It should be noted that there are concerns about excess basil exposure. Estragole, a suspect procarcinogen/mutagenic found in basil, raises questions about the balance between benefits and risks with the use of this and other spices (Muller et al. 1994).
Now, the majority of evidence points to the antimutagenic effects of basil outweighing the potential adverse effects associated with estragole-induced cell damage (Jeurissen et al. 2008).
17.4 - CARAWAY
Caraway (Carum carvi), also known as "meridian fennel" or "Persian cumin," is native to western Asia, Europe, and northern Africa.
The principal agents in caraway oil are believed to be carvone or p-mentha-1,8-dien-2-one and limonene or p-mentha-1,8-diene, the precursors of carvone and anethofuran (Zheng, Kenney, and Lam 1992). Although caraway appears to be a potent antioxidant in vitro, it has not been adequately examined in humans.
Recently, Kapoor et al. (2010) showed that caraway essential oil and oleoresins were progressively effectively with dose as antioxidants and more effective than commercial butylated hydroxyanisole and butylated hydroxytoluene.
Caraway oil and its ethanol oleoresin showed better reductive power than the other oleoresins. The scavenging and reducing power against the diphenylpicrylhydrazyl (DPPH) radicals that caraway oil and oleoresins provide may be associated with their ability to donate hydrogen and the presence of reductones.
Mazaki et al. (2006) examined the effect of caraway seed extract on mutagenesis induced by N-methyl- N-nitro-N′-nitrosoguanidine (MNNG) in S. typhimurium strains that are deficient in DNA MGMT. Their results indicated that caraway does not directly inactivate MNNG and O 6-methylguanine-DNA MGMT may be involved in the response.
Animal models have also been used to explore the anticancer potential of caraway in sites ranging from colon to skin cancers.
Schwaireb (1993) examined dietary caraway oil for its effects on skin tumors induced by DMBA and croton oil in female BALB/c mice. Mice fed with a diet containing 3% caraway oil for 23 weeks from the beginning of tumor promotion decreased the number of mice with papillomas (p < .001), the number of papillomas per mouse (p < .0001), and the average papilloma volume (p < .0001).
The number of carcinomas in those animals provided with caraway oil were significantly less than in the controls (Schwaireb 1993). Deeptha et al. (2006) examined the effects of oral caraway (30, 60, and 90 mg/kg body weight per day for 15 weeks) on aberrant crypt foci in male Wistar rats treated with the carcinogen 1,2-dimethylhydrazine.
Aberrant foci are early morphological events that represent an important step in colon cancer progression. Treatment of rats with 60 mg/kg body weight of caraway decreased carcinogen-induced aberrant crypt foci, indicators of oxidative stress, and fecal bacterial enzyme activity.
Induction of GST by anticarcinogenic compounds is an important mechanism by which several spices, including caraway, may promote carcinogen detoxification and thereby lower cancer risk. Zheng, Kenney, and Lam (1992) reported that activity of the detoxifying enzyme GST in the liver increased markedly after gavage treatment with 20 mg carvone or limonene in A/J mice.
Carvone was also found to increase the GST activity in the forestomach by about 80% (p < .05), more than double the GST activity in the large intestinal mucosa (p < .05), and more than triple the GST activity in the small intestinal mucosa (p < .005).
Carvone also increased glutathione (GSH) in the lung (p < .005) and in the small (p < .05) and large intestinal mucosa (p < .05).
Caraway may also influence carcinogen activation by its ability to modify carcinogen bioactivation. Polycyclic aromatic hydrocarbons and halogenated aromatic compounds such as 2,3,7,8-tetrodibenzo-p-dioxin (TCDD) are bioactivated by the xenobiotic-metabolizing CYP genes to form reactive metabolites that bind to DNA. Naderi-Kalali et al. (2005) reported that caraway extracts were effective in inhibiting the induction of CYP1A1 and CYP1A1-related RNA in rat hepatoma (H4IIE) cells.
Caraway extracts >0.13 μM significantly inhibited CYP1A1 induction, as measured by the 2,3,7-ethoxyresorufin O-deethylase assay, with roughly a tenfold suppression in enzyme activity observed at concentrations of 1.3 and 13 μM, inhibiting TCDD-dependent induction by 50%-90%, depending on the solvent used (Naderi-Kalali et al. 2005).
Overall, changes in both phase I and II enzymes are consistent with the ability of caraway and its active constituent to lower chemically induced cancers.
The importance of caraway and its isolated components in drug detoxification mechanisms in humans remains largely unexplored.
17.5 - CARDAMOM
Cardamom refers to herbs within the Elettaria (green) and Amomum (black) genera of the ginger family Zingiberaceae.
Cardamom is a common ingredient used in Indian cooking and in various parts of Europe. As with many spices, cardamom has been demonstrated to have antioxidant properties. Kikuzaki, Kawai, and Nakatani (2001) examined extracts from black cardamom (Amomum subulatum) for their ability to scavenge radicals.
The ethyl acetate-soluble fraction, containing several phenolic compounds (protocatechualdehyde, protocatechuic acid, 1,7-bis(3,4-dihydroxyphenyl)hepta- 4E,6E-dien-3-one, and 2,3,7-trihydroxy-5-(3,4-dihydroxy-E-styryl)-6,7,8,9-tetrahydro-5H-benzocycloheptene), scavenged about 90% of DPPH radicals when provided at 100 μg/mL.
Interestingly, at lower concentrations, its radical scavenging activity was comparable to that of α-tocopherol (Kikuzaki, Kawai, and Nakatani 2001).
Feeding male albino Wistar rats with a high-fat diet supplemented with 10% black cardamom seed powder for 90 days was found to lower 2-thiobarbituric acid reactive substances (TBARS) by 28% (p < .05) in heart tissue (Dhuley 1999).
In addition, cardamom caused significant increases (p < .05) in several antioxidant enzymes including catalase, superoxide dismutase, and GST in both liver and heart compared to controls (Dhuley 1999).
The ability of cardamom to inhibit chemical carcinogenesis was shown by Banerjee et al. (1994), who observed cardamom oil feeding (10 μL daily for 2 weeks) caused a significant decrease in liver CYP content in Swiss albino mice (p < .05). A 30% increase in GST activity (p < .05) and sulfhydryl levels (p < .05) in the liver also accompanied the cardamom oil treatment.
These observations suggest that intake of cardamom oil affects the enzymes associated with xenobiotic metabolism and may therefore have benefits as a deterrent to cancer (Banerjee et al. 1994).
Cardamom has also been demonstrated to decrease azoxymethane-induced colon carcinogenesis by virtue of its anti-inflammatory, antiproliferative, and proapoptotic activities. Providing aqueous cardamom suspensions can enhance detoxifying enzyme (GST activity) and decrease lipid peroxidation (Bhattacharjee, Rana, and Sengupta 2007).
Recently, cardamom aqueous extracts (1, 10, 50, and 100 mg/mL) were reported to significantly enhance splenocyte proliferation in a dose-dependent manner, especially when combined with black pepper (Majdalawieh and Carr 2010).
While the effects of cardamom and black pepper were the opposite on T helper-1 and -2 cytokine release by splenocytes, the presence of both spices significantly enhanced the cytotoxic activity of natural killer cells against YAC-1 lymphoma cells.
These findings provide evidence that cardamom may have anticancer benefits by modifying immunocompetence.
17.6 - CINNAMON
Cinnamon is a spice obtained from the bark of an evergreen tree belonging to the Lauraceae family.
Major constituents in cinnamon include cinnamaldehyde, eugenol, terpinene, α-pinene, carvacrol, linalool, safrole, benzyl benzoate, and coumarin (Tabak, Armon, and Neeman 1999).
Cinnamon is widely used in traditional Chinese medicine. Several studies have examined its antioxidant properties. When inbred male albino Wistar rats were fed a high-fat diet with 10% cinnamon bark powder (Cinnamomum verum) for 90 days, oxidative stress was substantially decreased, as evident by a reduction in TBARS, a biomarker of free radical production (Dhuley 1999).
Providing rats with cinnamon bark powder significantly increased several antioxidant-related enzymes, including catalase, superoxide dismutase, and GST in both liver and heart tissue, compared to controls. Glucose-6-phosphate dehydrogenase and GPx were also significantly increased (p < .05) in rats fed with cinnamon bark powder.
These enzymes help maintain GSH levels, essential for cellular integrity and protection against oxidative damage from free radicals (Dhuley 1999).
The ability of cinnamon extracts to suppress the in vitro growth of H. pylori, a recognized risk factor for gastric cancer, gastric mucosa-associated lymphoid tissue lymphoma, and possibly pancreatic cancer, has stirred considerable interest in the potential use of this spice to suppress human cancers (Farinha and Gascoyne 2005; Eslick 2006).
However, a pilot study involving 15 subjects given a cinnamon extract (80 mg/day) for 4 weeks was not effective in those infected with H. pylori (Nir et al. 2000). In this study, the colonization rate of H. pylori was measured by urea breath tests (UBTs).
Although a decline in H. pylori counts was observed in six patients with exceptionally high counts according to their first UBT, the study did not demonstrate a decrease in colonization overall, and in some individuals increased colony counts occurred. The lack of success of cinnamon as a single treatment regimen against H. pylori is not terribly surprising given the consistent failure of single-agent antibiotic trials.
Additional trials using higher cinnamon amounts and possibly in combination with other agents may be warranted to truly evaluate the effects of this spice (Nir et al. 2000).
Tabak et al. (1996) examined several spices for their ability to inhibit H. pylori. Cinnamon and thyme were found to be the most potent inhibitors of H. pylori growth and urease activity. Tabak, Armon, and Neeman (1999) reported cinnamon’s antibacterial activity against seven clinical isolates of H. pylori and the antiurease activity of two different cinnamon extracts (methylene chloride and ethanol) and their chemical constituents.
They found that adding 100 μg cinnamon per disk produced an inhibition zone of approximately 80 mm wide, which was greater than the inhibition zones produced by several antibiotics (10 μg ampicillin, 30 μg tetracycline, 15 μg erythromycin, 30 μg nalidixic acid, and 25 μg co-trimoxazole). Although a concentration of 25 μg/mL completely inhibited four H. pylori strains, 50 μg/mL was the minimum inhibitory concentration for all seven strains.
In a liquid medium, cinnamon extract began to inhibit H. pylori at a concentration >3 μg/ mL and peaked at a concentration >12 μg/mL, and a similar pattern of inhibition was observed with urease.
The efficiency of cinnamon extracts in inhibiting H. pylori in a liquid medium and its resistance to low pH may enhance its effects in an environment like the human stomach. The antibacterial effects of cinnamon extract may be due to cinnamaldehyde.
Adding 200 μg cinnamaldehyde per disk produced an inhibition zone >90 mm, while eugenol (2000 μg/disk) produced an inhibition zone of 68 mm, and carvacrol (2000 μg/disk) produced an inhibition zone of 66 mm; Tabak, Armon, and Neeman 1999).
Cao, Urban, and Anderson (2008) studied the role of polyphenolic polymers from commercial cinnamon extract in immune regulation using mouse RAW264.7 macrophages.
The authors examined whether cinnamon polyphenol extract (CPE) regulated immune function by affecting expression levels of genes that code for tristetraprolin (TTP/zinc finger protein 36), proinflammatory cytokines, and glucose transporter (GLUT) family proteins, and they compared these effects with that of insulin and lipopolysaccharide.
Because TTP downregulates proinflammatory cytokines, it has the potential for use in the prevention and treatment of inflammation-related diseases.
In this study, CPE rapidly increased TTP mRNA and protein levels in mouse RAW264.7 macrophages after 30 minutes of treatment, and twofold increase in expression was sustained throughout the 4 hours of treatment.
CPE also increased the mRNAs coding for proinflammatory cytokines, such as tumor necrosis factor α, cyclooxygenase-2, and interleukin 6, although the TTP levels were 6- to 3000-fold higher than the proinflammatory cytokine mRNA molecules in the same cells (Cao and Prior 1998).
In mammals, glucose is a critically important molecule in the host immune response to injury and infection, which is facilitated by GLUT family proteins, and based on this study, cinnamon increases GLUT expression.
Vascular endothelial growth factor (VEGF) is a critical factor in the induction of angiogenesis. Unfortunately, the side effects associated with most anti-VEGF drugs limit their use, and thus, the use of naturally occurring dietary inhibitors derived from diets has great appeal.
A water-based extract from cinnamon is a promising effective agent because it directly inhibits kinase activity of purified VEGFR2 as well as mitogen-activated protein kinase- and Stat3-mediated signaling pathway in endothelial cells (ECs; Lu et al. 2010).
Impressively, the extract was found to inhibit VEGF-induced EC proliferation, migration and tube formation in vitro, sprout formation from aortic ring ex vivo, and tumor-induced blood vessel formation in vivo. Polyphenols in the extract appear to be responsible for the response; cinnamaldehyde is observed to have little effect on VEGFR2 kinase activity (Lu et al. 2010).
While the ability of cinnamon to influence angiogenesis is intriguing, additional studies are warranted.
17.7 - CLOVE
Clove is derived from flower buds of the Eugenia caryophyllata tree. Several bioactive components are found in clove, including tannins, terpenoids, eugenol, and acetyleugenol (Kluth et al. 2007).
Cloves are native to Indonesia and are used in cuisines throughout the world. While no studies have been conducted in humans to date to evaluate use of cloves in cancer prevention, a few studies conducted in mice suggest its effectiveness, especially in modifying cellular detoxification processes. Feeding 40 mg clove per gram of diet to mice resulted in an increase in GST activity compared to those that were not fed the spice.
The physiological significance of these findings remains unclear because the increase was approximately 2% above normal in the liver (not significant), 18% in the stomach (p < .05), and 33% in the esophagus (p < .05; Aruna and Sivaramakrishnan 1990). An increase in GSH concentrations in the stomach also occurred (p < .05), suggesting detoxification of clove constituents in the stomach (Aruna and Sivaramakrishnan 1990).
In another study, feeding clove (0.5%, 1%, and 2%) to mice for varying durations (10, 20, and 30 days) modified several phase II enzymes associated with carcinogen bioactivation (Kumari 1991).
A dose-dependent response was observed for several detoxification enzymes. After 20 days, all exposures resulted in a significant increase in GST and cytochrome b5 (Kumari 1991).
DT-diaphorase was significantly elevated in those that were provided with 1% or 2% clove after 30 days (p < .0005) compared to controls. A significant decrease in CYP activity was observed in those provided with any clove after 30 days.
No changes were observed in the activity levels of aryl hydrocarbon hydroxylase in response to the clove administration. Malondialdehyde (MDA) formation was measured to monitor radiation-induced lipid peroxidation, and both the concentration of clove provided and the duration of exposure affected the results. Diets containing 2% clove for at least 20 days, or any concentration administered for 30 days, significantly decreased MDA (Kumari 1991).
Eugenol, an allyl chain-substituted guaiacol, may be responsible, at least in part, for the induction of the phase II enzymes (Han et al. 2007) and/or serve as an antioxidant (Rajakumar and Rao 1993; Nagababu and Lakshmaiah 1994).
Changes in phase I and II enzymes may account for the ability of eugenol to serve as an antimutagen (Miyazawa and Hisama 2003) and to inhibit carcinogen-induced genotoxicity (Han et al. 2007).
Kluth et al. (2007) examined the influence of several spice extracts on phase I and II enzymes in cultured human liver carcinoma and human colon adenocarcinoma cells, and they suggested a shift in the nuclear transcription factor Nrf2 was responsible for the induction.
Evidence also exists that clove extracts might interfere with β-catenin activity and thereby decrease colon carcinogenesis, but further studies are needed on this (Aggarwal 2010).
Similar to allspice, clove contains high amounts of eugenol. However, this compound cannot serve to increase gastrointestinal GPx promoter activity, suggesting other compounds in clove may account for its biological activity (Kluth et al. 2007).
Overall, the findings to date suggest that tissues adapt to exposures to one or more constituents in cloves. In doing so, clove may improve the ability of selected tissues to handle foreign compounds that might lead to the initiation of carcinogenesis.
Based on findings to date, additional clinical studies are warranted to determine the ability of clove to influence drug detoxification pathways.
17.8 - CORIANDER
Coriander (Coriandrum sativum) is an herb in the family Apiaceae and is native to southern Europe and northern Africa to southwestern Asia.
Although all parts of the plant are edible, its fresh leaves and dried seeds are most frequently used in cooking. Coriander is a common ingredient in many foods throughout the world. One of its principal constituents is linalool. Several animal studies provide evidence that coriander seeds can promote the hepatic antioxidant system.
Feeding a 10% coriander seed diet to male Wistar rats for 12 weeks decreased the ability of hexachlorocyclo-hexane, an organochlorine insecticide, to promote lipid peroxidation (Aruna and Sivaramakrishnan 1990; Anilakumar, Nagaraj, and Santhanam 2001).
Coriander can also influence foreign compound metabolism.
Feeding Swiss mice with 160-mg coriander seeds per gram diet resulted in GST induction ranging from 20% to 37%, depending on the tissue examined. In another study, Banerjee et al. (1994) observed roughly a doubling in GST activity in Swiss albino mice that were provided with diets containing coriander oil (10 μL coriander oil daily for 2 weeks).
No significant changes were observed in CYP or aryl hydrocarbon hydroxylase. Although relatively few studies focus on coriander for its anticancer properties, those that are available suggest coriander may be important (Esiyok, Otles, and Akcicek 2004).
17.9 - CUMIN
Cumin (Cuminum cyminum) is a flowering plant in the family Apiaceae and is native to the eastern Mediterranean region and India.
Thymoquinone (TQ) is the most abundant component of black cumin seed oil. TQ has been reported to exhibit antioxidant, antimicrobial, anti-inflammatory, and chemopreventive properties (Allahghadri et al. 2010; Nader, el-Agamy, and Suddek 2010) and to ameliorate B(a)P-induced carcinogenesis in the forestomach.
Swiss mice that were fed 160-mg cumin seeds per gram of diet and injected with B(a)P to induce chromosome aberrations were able to suppress the aberrations by 83% compared to controls (Aruna and Sivaramakrishnan 1990).
Part of this response may be due to cumin’s ability to influence phase II enzymes. In another study by Banerjee et al., feeding Swiss mice with 10-μL cumin oil daily for 2 weeks caused a 13% increase in GST levels (p < .1).
No statistically significant changes were observed in CYP activity, aryl hydrocarbon hydroxylase levels, or sulfhydryl levels in the liver compared to the control group, and thus, the first, and possibly most important, change may be an elevation in GST activity (Banerjee et al. 1994). Aruna et al. (2005) studied male albino Wistar rats to determine the effect of cumin seeds (0.25 g cumin/kg body weight) on oxidative stress induced by alcohol and heated sunflower oil, a source of polyunsaturated fatty acids.
The rats’ antioxidant status was determined to be near normal when cumin was consumed with alcohol and preheated oil, potentially due to cumin’s antioxidant and detoxification properties (Aruna, Rukkumani, and Menon 2005).
Considerable evidence points to the ability of TQ to suppress tumor cell proliferation, including colorectal carcinoma, breast adenocarcinoma, osteosarcoma, ovarian carcinoma, myeloblastic leukemia, and pancreatic carcinoma (Gali-Muhtasib, Roessner, and Schneider-Stock 2006).
Normal cells appear to be slightly resistant to TQ (Worthen, Ghosheh, and Crooks 1998). Several mechanisms may explain the ability of TQ to bring about a change in cell division in neoplastic cells, including downregulation in Bcl-xL, cyclin D1, and VEGF (Aggarwal et al. 2008).
Considerable evidence points to the ability of TQ to induce free radical formation in tumor cells. Thus, the biological response in tumor cells (pro-oxidants) may be different than that in normal cells (antioxidant; Koka et al. 2010).
TQ has also been found to be effective in inhibiting human umbilical vein EC migration, invasion, and tube formation, suggesting its role in angiogenesis (Yi et al. 2008). TQ (6 mg/kg/day) was also found to prevent tumor angiogenesis in a xenograft human prostate cancer (PC-3) model (Yi et al. 2008).
The multitude of effects caused by cumin serves as justification for its continued examination as a spice with widespread potential for health promotion.
17.10 - DILL
Dill (Anethum graveolens) is a relatively short-lived perennial spice.
Dill is an herb that in effect has two components that are dependent on the seasons. In the early spring, dill is used for its leaves and in the autumn for its seeds. The principal constituents of dill weed oil are anethofuran or 3,6-dimethyl-2,3,3a,4,5,7a-hydroxobenzofuran, and carvone or p-mentha-1,8-dien-2-one (Zheng, Kenney, and Lam 1992).
As with other spices, there is evidence that dill promotes drug detoxification mechanisms.
Providing 20 mg each of carvone and anethofuran by gavage once every 2 days for a total of three doses increased GST activity in A/J mice (Zheng, Kenney, and Lam 1992). The response depended on the agent and the tissue examined.
Anethofuran more than doubled the activity of the detoxifying enzyme GST in the liver (p < .005) and forestomach (p < .005), and carvone increased GST activity 78% in the forestomach (p < .05) and increased GST activity more than twofold in the liver and large intestinal mucosa (p < .05) and more than threefold in the small intestinal mucosa (p < .005; Zheng, Kenney, and Lam 1992).
Because GSH helps maintain cellular oxidation-reduction balance and protects cells against free-radical species, the combination of increased GST and GSH levels results may be particularly helpful in detoxifying foreign compounds, including carcinogens.
17.11 - GARLIC
Garlic (Allium sativum) is a member of the onion family Alliaceae.
Garlic has been used throughout history for both its culinary and medicinal properties. Garlic’s distinctive characteristics arise from sulfur, which constitutes almost 1% of its dry weight. The primary sulfur-containing constituents are γ-glutamyl-S-alk(en)yl-L-cysteines and S-alk(en)yl-L-cysteine sulfoxides.
Considerable variation in the S-alk(en)ylcysteine sulfoxide content can occur; alliin (S-allylcysteine sulfoxide) is the largest contributor. Alliin concentrations can increase during storage because of the transformation of γ-glutamylcysteines.
Although garlic does not typically serve as a major source of essential nutrients, it may contribute to several dietary factors with potential health benefits, including the presence of oligosaccharides, arginine-rich proteins and, depending on soil and growing conditions, selenium and flavonoids.
Preclinical models provide rather compelling evidence that garlic and its associated components can lower the incidence of breast, colon, skin, uterine, esophagus, and lung cancers. However, evidence in human investigations is less compelling.
Suppression of nitrosamine formation continues to surface as one of the most likely mechanisms by which garlic retards cancer. The ability of S-allyl cysteine (SAC) and its nonallyl analog S-propyl cysteine to retard N-nitroso compounds formation, but not diallyl disulfide (DADS), dipropyl disulfide, and diallyl sulfide (DAS), reveal the critical role that the cysteine residue plays in inhibition (Milner 2001).
Some of the most compelling evidence in humans comes from studies by Mei et al. (1989) demonstrating that ingesting 5 g/day of garlic blocked the enhanced urinary excretion of nitrosoproline resulting from exaggerated nitrate and proline intake.
More recent evidence suggests as little as 1 g of garlic may be sufficient to suppress nitroproline formation (Cope et al. 2009).
The ability of garlic to inhibit tumors due to different cancer-inducing agents and in different tissues indicates that a generalized cellular event is likely responsible for the change in tumor incidence and that the response is highly dependent on environmental or other types of biological insults. Because metabolic activation is required for many of these carcinogens, there is likelihood that either phase I or II enzymes are altered.
Interestingly, little change in CYP1A1, 1A2, 2B1, or 3A4 activities has been detected following treatment with garlic or related sulfur compounds.
However, this lack of responsiveness may relate to the amount and duration of exposure, the quantity of carcinogen administered, or the methods used to assess the cytochrome content or activity.
Wu et al. (2002), using immunoblot assays, found that the protein content of CYP1A1, 2B1, and 3A1 was increased by garlic oil and each of several isolated disulfide compounds. Their data demonstrated that the number of sulfur atoms in the allyl compound is inversely related to the depression in these cytochromes.
Several lipid- and water-soluble organosulfur compounds have been examined for their antiproliferative efficacy.
Some of the more commonly used lipid-soluble allyl sulfur compounds in tumorigenesis research are ajoene, DAS, DADS, and diallyl trisulfide (DATS). A breakdown of allicin appears to be necessary for achieving maximum tumor inhibition.
Earlier studies reported that lipid-soluble DAS, DADS, and DATS (100 μM) were more effective in suppressing canine tumor cell proliferation than isomolar water-soluble SAC, S-ethyl cysteine and S-propyl cysteine (Knowles and Milner 2001).
Undeniably, not all allyl sulfur compounds from garlic are equally effective in retarding tumor proliferation. Allyl sulfur compounds preferentially suppress neoplastic over non-neoplastic cells (Sakamoto, Lawson, and Milner 1997).
S-allylmercaptocysteine (SAMC), DAS, and DADS have also been reported to increase the percentage of cells blocked within the G2/M phase. p34cdc2 kinase is a complex that governs the progression of cells from the G2 phase into the M phase of the cell cycle (Knowles and Milner 2001).
Using LNCaP and HCT-116 human cancer cells, Xiao, Zeng, and Singh (2009) demonstrated that checkpoint kinase 1-mediated mitotic arrest resulting from DATS is the key to apoptosis induction. It is becoming increasingly clear that the response to allyl sulfurs relates to their ability to form free radicals rather than to serve as an antioxidant (Antosiewicz et al. 2008).
Allyl sulfurs may bring about changes by influencing the genomic expression by affecting histone homeostasis.
Allyl mercaptan is a particularly potent inhibitor of histone deacetylase (HDAC; Nian et al. 2009). HDAC inhibition has the potential to derepress epigenetically silenced genes in cancer cells, leading to cell-cycle arrest and apoptosis.
Sp3 appears to have a role in driving p21 gene expression after HDAC inhibition by allyl sulfur compounds and coincides with cell-cycle arrest.
Alliin has been reported to influence angiogenesis. It causes a dose-dependent inhibition of fibroblast growth factor 2 (FGF-2)-induced human EC tube formation and angiogen- esis in the chick chorioallantoic membrane model (Mousa and Mousa 2005). Xiao et al. (2006) suggested the antiangiogenic characteristics of DATS relate to its ability to downregulate VEGF secretion and VEGF receptor-2 protein level and inactivation of Akt kinase.
However, while DATS was effective in decreasing prostate cancer multiplicity in the transgenic adenocarcinoma of the mouse prostate model, it did not appear to relate to a change in angiogenesis (Singh et al. 2008).
17.12 - GINGER
Ginger (Zingiber officinale) is a member of the Zingiberaceae family and is consumed widely not only as a spice but also as a medicinal agent (see also Chapter 7 on ginger). Other members of the family include turmeric and cardamom.
Ginger’s cultivation appears to have begun in South Asia and has now spread to various parts of the world. It is sometimes called "root ginger" to distinguish it from other products that share the name.
The principal constituents of ginger include [6]-gingerol, [6]-paradol, [6]-shogaol (dehydration gingerols), and zingerone. Several studies have investigated ginger’s antioxidant properties (Chrubasik, Pittler, and Roufogalis 2005).
Gingerol has also been shown to decrease intracellular ROS formation in human keratinocyte cells (Kim et al. 2007), inhibit angiogenesis in human ECs, and limit nitrogen oxide synthase expression and epidermal growth factor-induced cell transformation and AP-1 transcriptional complexes in JB6 cells (Bode et al. 2001; Ippoushi et al. 2003; Davies et al. 2005; Kim et al. 2005).
Feeding NIN/Wistar rats a diet containing up to 0.5-5% ginger for 1 month significantly increased (p < .05) several liver antioxidant enzymes, including superoxide dismutase (76–141%), catalase (37–94%), and GPx (11–30%; Kota, Krishna, and Polasa 2008).
Lipid and protein oxidation was inhibited in rats consuming ginger, as evidenced by significant decreases (p < .05) in liver and kidney levels of MDA (35-59% and 27-59%, respectively) and carbonyl levels (23-36%), compared to controls (Kota, Krishna, and Polasa 2008).
Ippoushi et al. (2007) found that AIN-76 basal diets with 2% ginger decreased TBARS by 29% (p < .05) and suppressed 8-hydroxy-2′-deoxyguanosine (8-OHdG, a product of oxidative DNA damage) levels in Wistar rats.
TBARS was also significantly decreased (p < .001) in Wistar rats fed with diets supplemented with 1% ginger following exposure to lindane, a pesticide that is a global pollutant, (Ahmed et al. 2008).
Various animal models have been used to examine the role of ginger in cancer prevention. For example, Ihlaseh et al. (2006) exposed male Wistar rats to N-butyl-N-(4-hydroxybutyl)-nitrosamine (BNN) and uracil salt to induce tumors resembling human low-grade papillary urothelial neoplasia.
Rats fed with a basal diet supplemented with 1% ginger extract for 26 weeks had significantly fewer urothelial lesions compared to the controls or those fed with the diet with 0.5% ginger (p = .013; Ihlaseh et al. 2006).
However, ginger does not appear effective in all cases, as evidenced by the lack of protection against proliferative lesions in the bladders of Swiss mice fed with a 1% or 2% extract and exposed to BNN/N-methyl-N-nitrosourea (Bidinotto et al. 2006).
Induction of phase I and II activities may partially account for ginger’s anticarcinogenic actions. Banerjee et al. (1994) found that providing 10-μL ginger oil daily for 2 weeks to Swiss mice increased aryl hydrocarbon hydroxylase activity about 25% (p < .05) and increased GST by 60% (p < .01). No significant increase in GST induction was observed in Swiss mice fed with 160 mg ginger/gram diet (Aruna and Sivaramakrishnan 1990).
Inflammation is a significant risk factor for cancer, including prostate cancer. Mitogen-activated protein kinase phosphatase-5 (MKP5) is implicated as a proinflammatory inhibitor in innate and adaptive immune response in vivo (Zhang et al. 2004).
Providing [6]-gingerol upregulated MKP5 expression in normal prostate epithelial cells treated with 50 μM gingerol; likewise, it upregulated MKP5 expression in human prostate cancer cell lines (DU145, PC-3, LNCaP and LAPC-4; Nonn, Duong, and Peehl 2007).
Ginger extracts, more so than their individual components, have been shown to inhibit lipopolysaccharide-induced prostaglandin E2 (PGE2) production to an extent similar to that of indomethacin, a nonsteroidal anti-inflammatory drug.
Subfractions of ginger extract decreased LPS-induced COX-2 mRNA expression levels, although apparently not through the nuclear factor κB (NF-κβ) or activating protein 1 (AP-1) transcription factor pathways, because the ginger extracts did not inhibit TNF-α production (Lantz et al. 2007).
[6]-paradol, another active compound in ginger, is reported to induce apoptosis in human promyelocytic leukemia cells, JB6 cells, an oral squamous carcinoma cell line, and Jurkat human T-cell leukemia cells in a dosedependent manner (Huang, Ma, and Dong 1996; Lee and Surh 1998; Keum et al. 2002; Miyoshi et al. 2003). It is unclear whether [6]-paradol has molecular targets similar to [6]-gingerol.
Ginger also appears to have antitumorigenic properties. Several cell lines have been examined for their sensitivity to ginger. For example, alcoholic extracts of ginger inhibited tumor cell growth for Dalton’s lymphocytic ascites tumor cells and human lymphocytes at concentrations of 0.2-1 mg/mL in vitro (Unnikrishnan and Kuttan 1988).
In a study of cytotoxic activities of several compounds in ginger against four tumor cell lines (A549, human lung cancer; SK-OV-3, human ovarian cancer; SK-MEL-2, human skin cancer; and HCT-15, human colon cancer), [6]-shogaol was the most potent (ED50: 1.05–1.76 μg/mL), and [4]-, [6]-, [8]-, and [10]-gingerol displayed moderate cytotoxicity (ED50: 4.92-30.05; Kim et al. 2008).
Adding [6]-gingerol (25 μM) has been reported to inhibit proliferation in rat ascites hepatoma cells AH109A and increase apoptosis at higher concentrations (50 μM; Yagihashi, Miura, and Yagasaki 2008). Likewise, adding [6]-shogoal (60 μM) to COLO295 cells has been reported to increase the expression of GADD153, a gene that promotes apoptosis (Chen et al. 2007).
[6]-shogaol (>50 μM) also provokes DNA damage and apoptosis through an oxidative stressmediated caspase-dependent pathway (Chen et al. 2007).
Similarly, incubation of HEp-2 cells with ginger (250 μg/mL, 500 μg/mL, or 1000 μg/mL) resulted in a dose-dependent decrease in nitrite generation, increased production of superoxide, and decreased GSH levels compared to untreated cells, indicating ginger-induced apoptosis through the generation of ROS (Chen et al. 2007).
Ginger is also recognized for its potential usefulness to reduce nausea.
To determine whether ginger had antiemetic effects in cisplatin-induced emesis, Manusirivithaya et al. (2004) conducted a randomized, double-blinded, crossover study in 48 gynecologic cancer patients. The addition of ginger (1 g/day) to a standard antiemetic regimen has no advantage in reducing nausea or vomiting in the acute phase of cisplatin-induced emesis.
In the delayed phase, ginger and metoclopramide have no statistically significant difference in efficacy (Manusirivithaya et al. 2004). In another study, 1000 mg of ginger was compared to 20-mg intravenous (IV) metoclopramide, and to 4-mg IV ondansetron in controlling nausea in patients receiving cyclophosphamide chemotherapy.
Ginger was determined to be as effective as metoclopramide, but neither was as effective as ondansetron (Sontakke, Thawani, and Naik 2003).
Overall, while the anticancer findings of ginger are intriguing and several processes may be associated with the observed responses, additional studies are needed to clarify the underlying mechanisms and to determine overall benefits to humans (Pan et al. 2008).
17.13 - ROSEMARY
Rosemary (Rosmarinus officinalis) is a woody herb with fragrant needle-like leaves.
Rosemary is native to the Mediterranean region and possesses a bitter, astringent taste and highly aromatic characteristics that complement a wide variety of foods. Rosemary is a member of the family Lamiaceae, and it contains a number of potentially biologically active compounds, including antioxidants such as carnosic acid and rosmarinic acid.
Other bioactive compounds include camphor (up to 20% in dry rosemary leaves), caffeic acid, ursolic acid, betulinic acid, rosmaridiphenol, and rosmanol.
Due to its high antioxidant activity, crude and refined extracts of rosemary are now widely available commercially (Ho et al. 2000). While the data are difficult to interpret, when rosemary is added along with other herbs to a balsamic vinegar preparation used in soups and salads, it appears to provide protection again oxidative stress in humans (Dragan et al. 2007).
Considerable evidence also suggests that rosemary extracts, or its isolated components, can retard chemically induced cancers. For example, topical application of a rosemary extract has been reported to block the initiation and promotion phases of B(a)P- and DMBA-mediated skin tumorigenesis (Huang et al. 1994).
Likewise, topical application of pure carnosol and ursolic acid also inhibited 12-0-tetradecanoylphorbol 13-acetate (TPA)-induced skin tumor promotion in DMBA-initiated mice (Huang et al. 1994).
Adding rosemary or carnosol has also been shown to retard DMBA-induced mammary cancer in rats (Singletary, MacDonald, and Wallig 1996).
The depression in tumors may occur because of a change in the types and amounts of DMBA adducts bound to DNA (Amagase et al. 1996). Although not extensively studied, such evidence suggests the ability of rosemary to influence drug-metabolizing enzymes.
Rosemary extracts and the active compounds carnosic acid and rosmarinic acid have been found to inhibit the proliferation of various human cancer cell lines, including,
Part of the antitumorigenic properties associated with rosemary may relate to a decrease in TNF-α-induced ROS generation and NF-κB activation, and thus enhanced TNF-α-induced apoptosis (Moon et al. 2010).
Carnosol was the most effective in reducing tumor proliferation. Carnosol is also known to induce apoptotic cell death in high-risk pre-B acute lymphoblastic leukemia (ALL; Dorrie, Sapala, and Zunino 2001). At least part of this response may relate to a decrease in Bcl-2.
Although carnosol may be effective, it may also interfere with the actions of some other antitumor agents. Zunino and Storms (2009) reported that carnosol decreased the percentage of cell death in the pre-B ALL lines SEM, RS4;11, and REH when combined with cytarabine, methotrexate, or vincristine compared to these chemotherapeutic agents alone.
Overall, these data suggest that carnosol, and possibly other constituents in rosemary, may block the terminal apoptotic events induced by some chemotherapeutic drugs and therefore may decrease the effectiveness of some standard therapies for leukemia.
17.14 - SAFFRON
Saffron is a spice derived from the flower of the saffron crocus (Crocus sativus) plant native to Southwest Asia.
It has historically been the world’s most expensive spice per unit weight. Saffron imparts a bitter taste and hay-like fragrance to food. Saffron likely contains more than 150 volatile and aroma-yielding compounds.
A carotenoid, α-crocin, comprises >10% of dry saffron’s mass and is responsible for the rich golden-yellow hue created when saffron is added to food dishes. Picrocrocin, a bitter glucoside, is responsible for saffron’s flavor.
Significant information points to the ability of saffron to inhibit cancer (Abdullaev 2003).
Aqueous saffron preparations have been reported to inhibit chemically induced skin carcinogenesis (Das, Chakrabarty, and Das 2004). Both changes in carcinogen bioactivation and tumor proliferation appear to occur. Saffron infusion given orally either before or after DMBA treatment increased GST, GPx, catalase, and superoxide dismutase in liver (Das, Das, and Saha 2010).
Saffron and crocus also have significant antitumorigenic properties. Similar to other spices, they appear to suppress cell growth in neoplastic cells to a greater extent than in normal cells (Aung et al. 2007).
The ability of crocin to decrease cell viability occurs in a concentration- and time-dependent manner (Bakshi et al. 2009).
The response is not limited to cells in culture because pancreatic xenografts are also influenced by saffron (4 mg/kg diet for 30 days; Dhar et al. 2009). The effects of tumor suppression also have an impact on the longevity of the host.
A significant increase in the life span of Dalton’s lymphoma-bearing animals was found in those provided with saffron (Bakshi et al. 2009).
The mechanism by which saffron suppresses tumor proliferation has not been adequately explored, but a shift in caspases and an increase in Bax protein are possible (Mousavi et al. 2009). When a saffron extract (200-2000 μg/mL) was added to MCF-7 cells in culture, there was a marked decrease in cell viability as concentration and duration of exposure increased (IC50 of 400 ± 18.5 μg/mL after 48 hours).
Analysis of DNA fragmentation by flow cytometry revealed apoptotic cell death in these cells (Mousavi et al. 2009). Saffron-induced apoptosis was inhibited by pan-caspase inhibitors, indicating the importance of this process in determining the response.
17.15 - THYME
Thyme is another culinary and medicinal herb.
Today, common usage refers to any or all members of the plant genus Thymus, also of the Lamiaceae family. Several active agents are reported, including thymol, carvacrol, apigenin, luteolin, tannins, γ-terpinene, and other oils (Aydin, Basaran, and Basaran 2005; Kluth et al. 2007).
Feeding thyme leaves (0.5% or 2.0%) or its phenolic compounds, thymol and carvacrol (50–200 mg/kg), has been reported to enhance xenobiotic-metabolizing enzymes, including phase I enzymes such as 7-ethoxycoumarin O-deethylase and phase II enzymes such as GST and quinone reductase (Sasaki et al. 2005).
Admittedly, the isolated components were more effective than feeding the leaf. Kluth et al. (2007) examined the effects of thyme on enzyme induction in cultured human liver carcinoma cells and human colon adenocarcinoma cells.
They observed a thyme extract to activate CYP3A4 promoter via PXR and the GI-GPx promoter via the electrophile responsive element, thus providing potential clues about the mechanism by which thymol and carvacrol may influence phase I and II enzyme expression (Kluth et al. 2007).
The number of studies on genotoxic effects of thymol and carvacrol are limited, but contradictory. S. typhimurium tests have provided some, but not compelling, evidence that thyme is weakly mutagenic (Stammati et al. 1999).
In vivo, 0.25% thyme had no detectable effect on mouse embryo development (Domaracky et al. 2007).
In comet assays with human lymphocytes, thymol and carvacrol did not induce DNA strand breakage at concentrations lower than 50-100 μM, and were therefore considered safe for consumers (Undeger et al. 2009).
17.16 - CONCLUSION
Mounting evidence suggests that cancers are not an inevitable consequence of aging but are preventable diseases.
The evidence in this chapter suggests that spices may be factors in one’s diet that may lower cancer risk and affect tumor behavior. Spices have been consumed for centuries for a variety of purposes, such as flavoring agents, colorants, and preservatives.
This chapter only scratches the surface of the overall impact of herbs and spices since there are approximately 180 spices commonly being used for culinary purposes.
Without question, evidence exists that multiple processes, including proliferation, apoptosis, angiogenesis, and immunocompetence, can be influenced by one or more spices.
While the currently available data are intriguing, considerably more information is needed to determine who will benefit most from exaggerated intake of one or more spices, the effective exposures needed to bring about the desired outcome(s), and what interactions (both positive and negative) exist with other components of the diet or with medications that an individual may regularly consume.
REFERENCES
|